

Ситовидные трубки
Ситовидные трубки (решётчатые трубки, ситовидные сосуды) — проводящие элементы в телах высших растений, проводящие сахар и пластические питательные вещества, развитые преимущественно в лубяной части сосудисто-волокнистого пучка. Главной их функцией является транспортировка углеводов — например, из листьев в плоды и корни. Были открыты и названы германским биологом и исследователем леса Теодором Гартигом в1837 году.
Ситовидные трубки представляют однорядные тяжи вытянутых в длину клеток, толстые и различных цветов; их конечные стенки являются ситовидными пластинками, несущими ситовидные поля с многочисленными прободениями, которые изнутри выстланы каллозой. Простые пластинки обычно горизонтальные и содержат одно ситовидное поле (характерными примерами являются тыква и ясень), сложные же расположены под наклоном и имеют несколько ситовидных полей (к таким растениям относятся липа, виноград, пассифлора, рис). К каждому членику ситовидной трубки примыкает тяж узких сопровождающих клеток, которые физиологически связаны с ними[1]. При их развитии тонопласты в клетках постепенно разрушаются, вызывая смешивание цитоплазмы с клеточным соком; происходит дегенерация органелл и ядра клетки. Ввиду значительного количества цитоплазмы не считаются «настоящими» клетками, они не имеют клеточного ядра, рибосом и вакуоли.
У большинства растений ситовидные трубки функционируют не более года, но встречаются и исключения: у винограда они существуют 2 года, у липы — в течение нескольких лет, тогда как у некоторых пальм — несколько десятков. В конце вегетационного периода ситовидные прободения полностью закупориваются каллозой, откладывающейся также на обеих сторонах ситовидной пластинки, вследствие чего образуются мозолистые тела. Более не функционирующие ситовидные трубки и окружающие клетки со временем деформируются и облитерируются.
КАЛЛОЗА, каллёза, полисахарид, содержащийся в растениях и состоящий из остатков молекулы глюкозы, соединенных Р-1,3 связями в спиральную цепочку. Каллоза нерастворима в воде, хорошо флюоресцирует в люминесцентном микроскопе. У виноградного растения каллоза образуется как в вегетативных, так и в генеративных органах. Она выстилает канальцы ситовидных пластинок флоэмы осенью 1-го года их образования, весной, гидролизуясь, рассасывается. Осенью 2-го года каллоза вновь закупоривает канальцы, а на 3-й год выводит ситовидные трубки из строя. На стенках клеток паренхимы пораженных органов каллоза способствует образованию каллуса. В клетках фиброзного слоя пыльника откладывающаяся каллоза образует утолщения. В оболочках микро- и макроспороцитов (и соответственно микро- и макроспор) каллоза играет роль изолятора, а также предохраняет от обезвоживания и др. неблагоприятных условий, что имеет важное значение при спорогенезе. В оболочке растущих пыльцевых трубок, особенно в базальной части, откладывающаяся каллоза вызывает утолщения, часто — каллозные пробки.
20) Проводящие пучки растений
Обособленные тяжи проводящей системы, состоящие чаще из ксилемы и флоэмы , называют проводящими пучками. Первоначально они возникают из прокамбия . Вокруг пучков нередко формируется обкладка из живых или мертвых паренхимных клеток. Они могут быть полными, т.е. состоящими из флоэмы и ксилемы , и неполными, состоящими только из ксилемы или флоэмы. В тех случаях, когда часть прокамбия сохраняется и превращается затем в камбий , а пучок способен к вторичному утолщению, говорят об открытых пучках. Они встречаются у большинства двудольных и голосеменных . В закрытых пучках однодольных прокамбий полностью дифференцируется в проводящие ткани.
В зависимости от взаимного расположения флоэмы и ксилемы различают пучки нескольких типов. Чаще всего флоэма лежит по одну сторону от ксилемы. Такие пучки называют коллатеральными (открытые и закрытые). У части двудольных растений(из семейств пасленовых , вьюнковых , тыквенных и т д.) одна часть флоэмы располагается снаружи, а другая - с внутренней стороны ксилемы. Такой пучок называется биколлатеральным, a соответствующие участки флоэмы - наружной и внутренней флоэмой. Камбий находится между наружной флоэмой и ксилемой .
Встречаются также концентрические пучки, при этом либо флоэма окружает ксилему (центроксилемные пучки), либо, наоборот, ксилема окружает флоэму (центрофлоэмные). Центрофлоэмные пучки найдены в стеблях и корневищах ряда двудольных ( ревень ,щавель , бегония ) и однодольных (многие лилейные , осоковые ). Известны пучки промежуточные между закрытыми коллатеральными и центрофлоэмными. Центроксилемные пучки обычны для папоротников .
В центре молодых корней голосеменных и покрытосеменных , имеющих первичное строение, располагается проводящий пучок, получивший название радиального. Ксилема в таком пучке расходится лучами от центра, а флоэма располагается между лучами. Возникают эти пучки из прокамбия . В корнях двудольных и голосеменных между ксилемой и флоэмой сохраняется слой прокамбиальных клеток, которые позднее дифференцируются в камбий . Встречаются однолучевые (монархные), двулучевые (диархные), трехлучевые (триархные), четырехлучевые (тетрархные), пятилучевые (пентархные) и многолучевые (полиархные) радиальные лучи. Последние обычны у однодольных .
Ксилема и флоэма обычно расположены рядом, образуя слои, или так называемые проводящие пучки, представленные в растениях несколькими типами. В зависимости от расположения ксилемы и флоэмы относительно друг друга, различают следующие типы:
1. Коллатеральные (бокобочные), когда ксилема и флоэма располагаются бок о бок, т.е. на одном радиусе (рис. 19, А, В).
2. Биколлатеральные (дважды бокобочные пучки) - флоэма прилегает к ксилеме с обеих сторон. Наружный участок флоэмы более мощный (рис. 19, Б).
3. Концентрические :
а) амфивазальные - ксилема замкнутым кольцом окружает флоэму (рис. 19, Г );
б) амфикрибральные - флоэма окружает ксилему (рис. 19, Д).
4. Радиальные - ксилема расходится лучами от центра, а флоэма располагается между лучами (рис. 19, Е).
Проводящие пучки в зависимости по наличию или отсутствию в них камбия бывают открытые и закрытые. В открытых - между ксилемой и флоэмой есть камбий (рис. 19, А, Б). В закрытых - камбия нет (рис. 19, В).
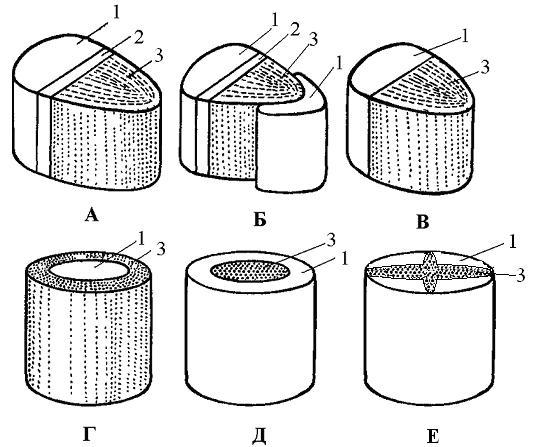
Рис. 19. Типы проводящих пучков:
А - открытый коллатеральный; Б - открытый биколлатеральный; В - закрытый коллатеральный; Г, Д - концентрические (Г - амфивазальный, Д - амфикрибральный); Е - радиальный.
1 - флоэма, 2 - камбий, 3 - ксилема.
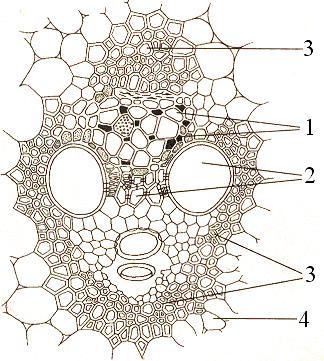
Рис. 20. Закрытый коллатеральный проводящий пучок на поперечном срезе стебля кукурузы:
1 - флоэма, 2 - ксилема, 3 - механическая обкладка пучка, 4 - основная паренхима стебля, окружающая пучок.
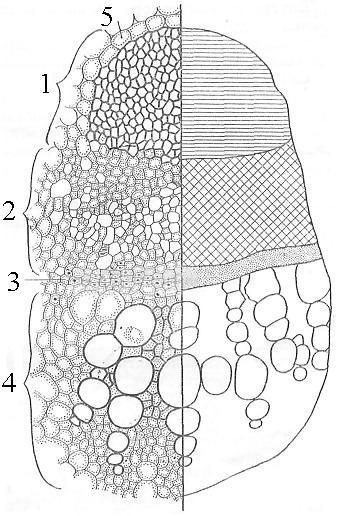
Рис. 21. Открытый коллатеральный проводящий пучок на поперечном срезе стебля подсолнечника (слева - детальный рисунок, справа - схематичный):
1 - склеренхима, 2 - флоэма, 3 - камбий, 4 - ксилема, 5 - основная паренхима стебля.
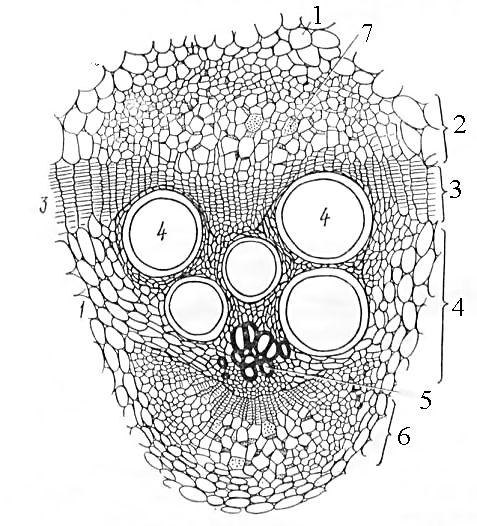
Рис. 22. Открытый биколлатеральный проводящий пучок на поперечном срезе стебля тыквы:
1 - основная паренхима стебля, 2 - наружная флоэма, 3 - камбий, 4 - вторичная ксилема, 5 - первичная ксилема, 6 - внутренняя флоэма, 7 - ситовидная пластинка.
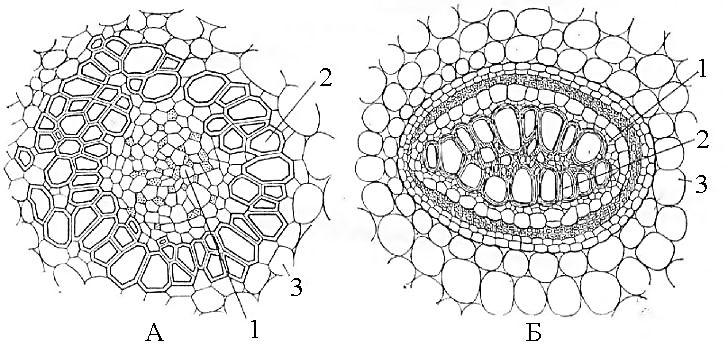
Рис. 23. Концентрические проводящие пучки:
А - амфивазальный пучок корневища ландыша; Б - амфикрибральный пучок корневища папоротника-орляка.
1 - флоэма, 2 - ксилема, 3 - основная паренхима стебля.
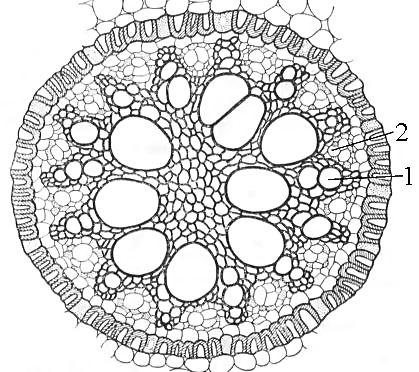
Рис. 24. Радиальный проводящий пучок корня ириса:
1 - луч ксилемы, 2 - участок флоэмы.
21) К выделительным (секреторным) тканям относятся разного рода структурные образования, способные активно выделять из растения или изолировать в его тканях продукты метаболизма и капельно-жидкую воду. Выделяемые наружу или накапливаемые внутри жидкие и твердые продукты метаболизма получили общее название секретов . Как правило, секреты (смесь терпеноидов ,полифенольные соединения , оксалат кальция ) относятся к продуктам вторичного метаболизма (обмена), но среди них встречаются и продукты первичного обмена .
Вторичные метаболиты представлены в растениях огромным числом индивидуальных соединений, хотя они образуются на немногих путях обмена веществ ( схема 1 ) и их биогенетическими предшественниками являются - мевалонат , ацетил-КоA (ацетил-коэнзим A) - сложное органическое вещество, молекулы которого участвуют в главнейших биохимических реакциях, идущих в живой клетке, коричная кислота и ряд белковых аминокислот.
В левой части схемы 1 указаны биогенетические предшественники основных классов соединений, относимых к продуктамвторичного метаболизма .
Три предшественника: мевалонат , ацетил-КоA и аминокислоты относятся к продуктам первичного обмена ; коричная кислота - продукт уже вторичного обмена, но она оказывается необходимой в ходе биохимического синтеза флавоноидов и различных более простых природных производных фенилпропана, обобщенно называемых фенилпропаноидами.
В правой колонке схемы перечислены главнейшие классы вторичных метаболитов (цифрами указано приблизительное число индивидуальных соединений, выделенное из различных групп организмов). Стрелками показана связь между теми или иными соединениями в ходе биохимических реакций. Сами биохимические реакции, в ходе которых одни соединения превращаются в другие, довольно разнообразны и включают множество стадий.
Элементы, или комплексы, выделительных тканей встречаются во всех органах. В зависимости от того, выделяют они вещества наружу или выделенные вещества остаются внутри растения, их делят на две группы: ткани внутренней и наружной секреции.
Клетки выделительных тканей по форме обычно паренхимные и тонкостенные. Они долго остаются живыми, выделяя секрет .Клетки-идиобласты по мере накопления большого количества секрета лишаются протопласта и стенки их нередко опробковевают. Синтез жидких секретов связывают с деятельностью внутриклеточных мембран и комплекса Гольджи .
Поскольку продукты вторичного метаболизма биологически активны и могут вызвать повреждение цитоплазмы , существуют механизмы, препятствующие этому. Один из них - перенос таких веществ в вакуоль или в свободное изолированное от цитоплазмы пространство клетки. Другой механизм - химическое превращение соединений до относительно безвредных, что, разумеется, не исключает их последующее выделение.
Прежде чем выделиться из цитоплазмы , где они синтезируются, секретируемые вещества преодолевают цитоплазматические мембраны - плазмалемму , если вещества выделяются в свободное пространство клетки, или тонопласт - при транспорте в вакуоль .
Эволюционно внутренние выделительные ткани возникли из ассимиляционных и запасающих , а наружные связаны с покровными тканями . Клетки, содержащие оксалат кальция , изначально выступают как ассимиляционные или запасающие и лишь позднее превращаются в выделительные.
Функции выделительных тканей растений существенно отличаются от функций выделительной системы животных. Образующиеся секреты нередко эффективно защищают растения от поедания животными, повреждения насекомыми или патогенными микроорганизмами.
Часто секреты , выступающие из мест поранения растений при искусственных или естественных повреждениях, играют роль бактерицидного пластыря (смолы, бальзамы). Выделяющиеся в цветках ароматические и сахаристые вещества ( нектар ) привлекают насекомых-опылителей. Наконец, накапливающиеся в разного рода вместилищах секретированные вещества могут вновь вовлекаться в процесс метаболизма и в этом случае выступают в роли запасных веществ. Клетки-идиобласты , особенно содержащиеоксалат кальция , приобретают значение мест длительного "захоронения" токсичных для растения веществ или веществ, полностью исключенных из метаболизма. Вещества, полностью исключающиеся из метаболизма, удаляются из растения при опадении листьев, слущивании корки и т.п. Это основной путь избавления от "шлаков".
Судьба секретирующих клеток различна. Иногда они остаются живыми длительное время. При этом секреция осуществляется путем пассивного или активного транспорта либо экзоцитоза . В иных случаях при секреции происходит повреждение клетки. Выделение наружу выработанного секрета сопровождается выбросом части цитоплазмы , но отделяется только безъядерная часть клетки. Наконец, известны случаи, когда клетка полностью дегенерирует и иногда вместе с выработанным ею продуктом выделяется в окружающую среду (например, солевые волоски некоторых галофитов и слизистые клетки корневого чехлика ).
Выделение капельно-жидкой воды характерно для многих растений и осуществляется через гидатоды .
Ткани внутренней секреции могут быть представлены отдельными клетками-идиобластами , вместилищами выделений , смоляными ходами , эфирномасляными каналами и млечниками ( рис. 49 ).
24) Ка́мбий (от лат. cambium — обмен, смена), образовательная ткань в стеблях и корнях преимущественно двудольных и голосеменныхрастений, дающая начало вторичным проводящим тканям и обеспечивающая рост их в толщину. Сезонные изменения активности камбия обусловливают образование годичных колец древесины.
образовательная ткань (Меристема) в корнях и стеблях преимущественно двудольных и голосеменных растений. К. расположен однорядным цилиндрическим слоем (на поперечном срезе — в виде кольца). В результате деятельности К. происходит прирост осевых органов в толщину. Он образует кнаружи вторичную флоэму (См. Флоэма) (луб) и кнутри — вторичную ксилему (См. Ксилема) (древесину). К. возникает из клеток прокамбия (См. Прокамбий), лежащих между первичными флоэмой и ксилемой. В образовании К. в корнях большое значение имеет Перицикл. В листьях К. нет, если же он есть, то его деятельность рано затухает. По форме клетки К. прозенхимные — удлинённо-заострённые (на концах скошены), таблитчатые, расположены по длине органа (рис. 1). Оболочки клеток К. мягкие, целлюлозные, имеют первичные поровые поля с плазмодесмами (См. Плазмодесмы)
Клетки К. делятся, видоизменяясь в клетки луба (кнаружи от К.) или древесины (кнутри от К.) (рис. 2). Обычно древесины образуется в несколько раз больше, чем луба. В результате деления некоторых клеток К. образуются мелкие клетки: это — лубо-древесинные лучи (рис. 3), производящие в одну сторону лубяную, в др.— древесинную часть луча. На осень и зиму деятельность К. прекращается (в умеренных широтах). Периодичность деятельности К. вызывает образование годичных колец (См. Годичные кольца). В зависимости от характера деления клеток К. различают ярусный и неярусный К.
У некоторых двудольных (например, у свёклы) в стеблях и корнях образуются последовательно кольцами к периферии дополнительные слои К. (поликамбиальность). О К., образующем пробку, см. Феллоген, Перидерма.
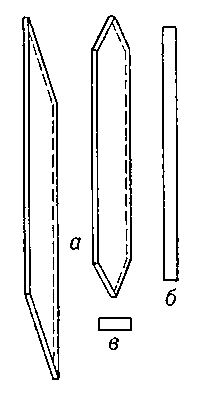
Рис. 1. Схема прозенхимной клетки камбия: а — продольный тангентальный разрез; б — продольный радиальный разрез; в — поперечный разрез.
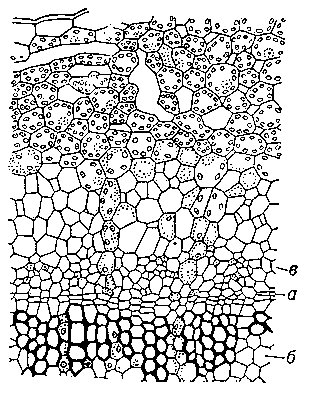
Рис. 2. Часть поперечного разреза стебля льна: а — камбий; б — древесина; в — луб.
Рис. 3. Клетки камбия, разделившиеся на клетки лубо-древесинного луча.
25) Хлоре́нхима, или хлорофилоносная паренхима — ассимиляционная ткань сосудистых растений, состоящая изпаренхимных клеток, вдоль тонких стенок которых одним слоем располагаются хлоропласты, не затеняя друг друга.
В некоторых руководствах ассимиляционную ткань рассматривают как разновидность основной паренхимы.