

Характеристика адренорецепторов
Адренорецепторами называют молекулы клетки, которые реагируют на нейромедиатор норадреналин или гормон адреналин. Впервые мысль о возможном существовании нескольких типов адренорецепторов была выдвинута английским фармакологом Ahlquistв 1948 г. В настоящее время адренорецепторы подразделяют на 2 класса:
-адренорецепторы – этот класс рецепторов опосредует сокращение гладких мышц под влиянием адреналина. Обнаружено 2 типа -адренорецепторов (1и2), каждый из которых имеет по крайней мере 3 подтипа.
-адренорецепторы – этот класс рецепторов опосредует расслабление гладких мышц под влиянием адреналина. Обнаружено 3 типа -адренорецепторов (1,2,3).
Все типы адренорецепторов являются семейством мембранных рецепторов, связанных с G-белками. Подробная их характеристика представлена в таблице 4.
Таблица 4. Сравнительная харктеристика адренорецепторов.
|
Тип |
Агонист |
Антагонист |
Локализация |
Функция |
Механизм |
|
1 |
A>NA>Iso Фенилэфрин |
Празозин |
Постсинаптические (на эффекторн. тканях) |
|
|
|
1А |
? |
Тамсулозин (+)-нигульдипин |
Миокард
Печень
Гладкие мышцы МПС |
Повышение сократимости, аритмия гликогенолиза, синтеза гликогена Сокращение |
Активация через Gq-белок фосфолипазы С, D и А2, а также L-типа Са2+-каналов |
|
1В |
? |
WB 4101 |
Гладкие мышцы ЖКТ |
Расслабление, уменьшение моторики и тонуса |
Активация Са2+-зависимых К+-каналов, гиперполяризация |
|
1D |
? |
? |
Гладкие мышцы сосудов кожи, слизистых, ЖКТ, почек и головного мозга |
Сокращение, повышение АД |
Как у 1А |
|
2 |
A>NA>Iso Клонидин |
Йохимбин |
Пресинаптические Внесинаптические |
|
|
|
2А |
Оксиметазолин |
? |
Постганглионарные волокна (пресинаптически) Тромбоциты Мышцы сосудов ЦНС |
Снижение секреции медиатора (норадреналина) Агрегация Сокращение, АД Седативное и анальгетическое действие |
Тормозят через Gi-белок аденилатциклазу, активируют К+-каналы, блокируют L- и N-тип Са2+-каналов |
|
2В |
? |
ARC 239 |
-клетки поджелудочной железы |
Снижение секреции инсулина | |
|
2C |
? |
ARC 239 |
|
| |
|
1 |
Iso>A=NA Добутамин |
Метопролол |
Постсинаптические Миокард
Клетки ЮГА |
автоматизма (ЧСС), проводимости и сократимости секреции ренина |
Активация через Gs-белок аденилатциклазы и L-типа Са2+-каналов |
|
2 |
Iso>A>>NA Сальбутамол |
Бутоксамин |
Пресинаптические Внесинаптические Гладкие мышцы (сосудов, дыхательных путей, ЖКТ, МПС) Скелетные мышцы Печень |
секреции NA
Расслабление
Гликогенолиз гликогенолиза синтеза гликогена | |
|
3 |
Iso=NA>A BRL 37344 |
SR 59230 CGP 20712A |
Внесинаптические Жировая ткань |
Липолиз |
Примечание: A – адреналин, NA – норадреналин, Iso – изопреналин, МПС – мочеполовая система.
1-адренорецепторыпосредствомGq-белка передают сигнал на несколько эффекторных систем:
Фосфолипазу С, которая гидролизует фосфатидилинозитол бифосфат (PIP2) до инозитов трифосфата (IP3) и диацилглицерола (DAG). МолекулыIP3вызывают выход ионов Са2+из внутриклеточного депо и активируют зависимые от Са2+ферменты (кальмодулин).DAG– обеспечивает активацию протеинкиназы С и фосфорилирование внутриклеточных белков, а также открывает Са2+-каналы мембраны. Под влиянием ионов Са2+и активного кальмодулина происходит дефосфорилирование киназы легких цепей миозина и она переходи в активную нефосфорилированную форму, при этом начинают фосфорилироваться легкие цепи миозина и запускается процесс сокращения гладкомышечных клеток (см. схему 6.).
Фосфолипазу А2, которая гидролизует фосфолипиды с выделение арахидоновой кислоты. В последующем арахидоновая кислота трансформируется в простагландины и лейкотриены.
Фосфолипазу D, которая гидролизует фосфатидилхолин до фосфатидной кислоты. Молекулы фосфатидной кислоты вызывают выделение ионов Са2+из депо, активируют АДФ-рибозилирующий фактор.
Показана возможность активации G-белками Са2+-каналов клетки.
2-адренорецепторыпосредствомGi-белка также передают сигнал на несколько эффекторных систем:
Gi-белок снижает активность аденилатциклазы и уменьшает синтез цАМФ в клетке. В итоге, активность зависимых от цАМФ протеинкиназ падает.
Через G0-белки тормозятся Са2+-каналыL- иN-типов.
-субъединицы G-белка активируют К+-каналы мембраны.
Относительно недавно было обнаружено, что -субъединицыGi-белка могут стимулировать митоген-активирующие протеинкиназы (МАРК), которые обеспечивают процессы деления и размножения стволовых клеток.
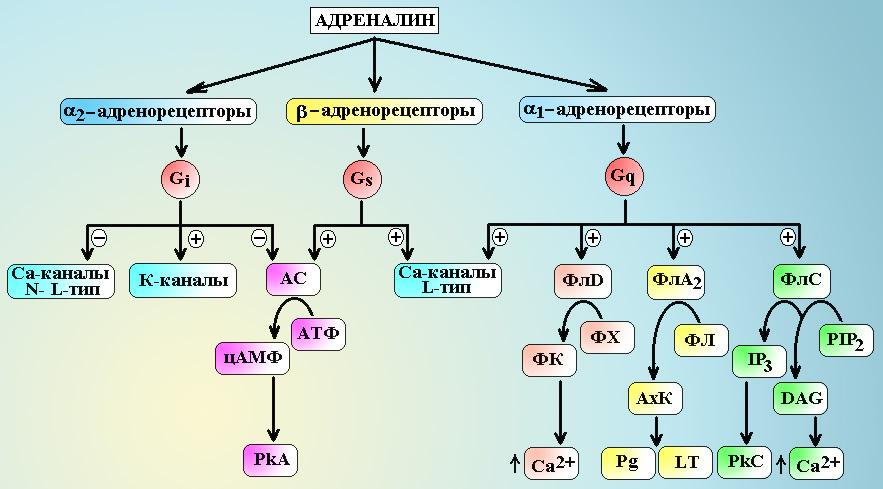
Схема 5. Передача сигнала с адренорецепторов. АС – аденилатциклаза, PkA – протеинкиназа А, PkC – протеинкиназа С, ФлС – фосфолипаза С, ФлА2 – фосфолипаза А2, ФлD – фосфолипаза D, ФХ – фосфатидилхолин, ФЛ – фосфолипиды, ФК – фосфатидная кислота, АхК – арахидоновая кислота, PIP2 – фосфатидилинозитол бифосфат, IP3 – инозитол трифосфат, DAG – диацилглицерол, Pg – простагландины, LT – лейкотриены.
-адренорецепторывсех типов реализуют свое действие черезGs-белки.-субъединицы этого белка активируют аденилатциклазу, которая обеспечивает синтез в клетке цАМФ из АТФ и активацию цАМФ зависимой протеинкиназы А.-субъединицыGs-белка активируют Са2+-каналыL-типа и т.н.maxi-K+-каналы. Под влиянием цАМФ-зависимой протеинкиназы А происходит фосфорилирование киназы легких цепей миозина и она переходит в неактивную форму, не способную фосфорилировать легкие цепи миозина. Процесс фосфорилирования легких цепей прекращается и гладкомышечная клетка расслабляется.