

Психогенетика_СГУ_1
.pdfСовременный Гуманитарный Университет
21

Современный Гуманитарный Университет
22
23 |
Гуманитарный Современный |
|
Университет |
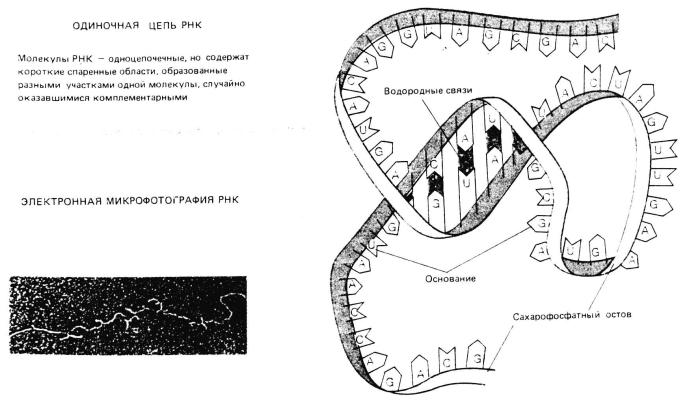
Рис. 3. Структура ДНК и РНК
Молекула ДНК (дезоксирибонуклеиновая кислота) представляет собой полимер, состоящий из остатка фосфорной кислоты, са хара дезоксирибозы и гетероциклических оснований: аденина, гу анина, тимина, цитозина. ДНК – это цепь нуклеотидов. Каждый нуклео тид состоит из азотистого основания, сахара дезоксирибозы и остатка ф осфорной кислоты. Между собой нуклеотиды соединены химической свя зью с остатками фосфорной кислоты. Все нуклеотиды имеют одинак овый сахар и остаток фосфорной кислоты. Две цепи ДНК соединены слабы ми водородными связями между азотистыми основаниями. Пары о снований соединяются между собой по так называемому принципу комплементарности. Аденин (А) всегда соединяется с тимино м (Т), а гуанин (Г) с цитозином (Ц). Двойные цепи ДНК закручены в спир аль, которая ограничена с двух сторон дезоксирибозой и фосфат ными группами. В результате в это пространство могут поместить ся только пары А-Т и Г-Ц, поскольку по своим размерам аденин и гуанин значи тельно больше тимина и цитозина. Следовательно, в это пространст во могут поместиться только сочетания оснований с большим и малым размерами. Другой причиной комплементарности является химическое строение азотистых оснований.
Знание молекулярных механизмов наследственности важно для понимания того, как наследственные факторы влияют на псих ику человека. Сами наследственные факторы не детерминируют п оведение, а определяют последовательность аминокислот в белках. Ме жду наследственными факторами и психическими характеристик ами имеются многочисленные контакты, происходящие на разных уровнях формирования организма. Раскрытие причинно-следственны х взаимоотношений между ними потребует больших усилий и мн ого времени.
Вследствие комплементарности двух нитей ДНК возможно то чное воспроизведение молекул ДНК. При репликации – процессе самовоспроизведения молекул ДНК – водородные связи межд у комплементарными нитями ДНК разрушаются, и для каждой из исходных нитей ДНК строится новая комплементарная нить. Существуе т образ такого процесса как застежка-молния, которая многократно расстегивается и застегивается без повреждения.
РНК - рибонуклеиновые кислоты – представляют собой полимеры, состоящие из остатка фосфорной кислоты, сахара рибозы, гетероциклических оснований: аденина, гуанина, урацила, ц итозина. Имеется несколько видов РНК, которые имеют разную структу ру и выполняют разные функции. Матричная (информационная) РНК (м- РНК) - молекула рибонуклеиновой кислоты, содержащая инфор мацию о последовательности аминокислот в белке, обеспечивает переписывание (транскрипцию) генетической информации с молекулы ДНК. Образно можно сказать, что ДНК – это чертеж, РНК – копия чертежа, которая используется в производстве. Транспортная РНК уч аствует в трансляции (переводе) последовательности нуклеотидов в м-РНК в
Современный Гуманитарный Университет
24
последовательность аминокислот в белковой цепи. Биосинт ез белка осуществляется органоидами белка - рибосомами, в которых имеются рибосомальная РНК.
При синтезе молекулы м-РНК одна из нитей ДНК служит матриц ей для построения комплементарной к ней молекуле РНК. Молеку лы м- РНК являются одноцепочечными и после окончания транскри пции они из ядра клетки выходят в цитоплазму и соединяются с рибос омами, образуя “фабрику” по синтезу белка.
Белковая молекула представляет собой цепочку аминокисл от, соединенных между собой пептидной связью. Всего имеется 2 0 аминокислот, из которых образуются все белки в организме. Синтез белков происходит со скоростью примерно 100 аминокислот в с екунду. Последовательность нуклеотидов в ДНК определяет послед овательность нуклеотидов в м-РНК, что, в свою очередь, определяет последовательность аминокислот в белке. Информация о стр оении белка – это информация, которая передается потомкам из по коления в поколение. Кодирование последовательности аминокислот последовательностью нуклеотидов и является кодированием наследственной информации.
При расшифровке генетического кода решается вопрос, как сочетание четырех азотистых оснований кодирует последо вательность из 20 аминокислот в белке. Если бы одно основание соответств овало одной аминокислоте, то белки состояли только из четырех а минокислот. Если бы два основания определяли положение аминокислоты в белке, то можно было бы кодировать только 16 аминокислот. Было уста новлено, что сочетание из трех оснований обеспечивает включение в сех 20 аминокислот в состав белка. В этом случае число возможных сочетаний из трех оснований (триплетов) равно 64. Поскольку число амин окислот меньше числа возможных триплетов, то некоторые аминокисл оты кодируются несколькими триплетами. Это явление получило название
вырожденности генетического кода. Некоторые триплеты, так называемые нонсенс-кодоны (УАГ, УАА, УГА), служат сигналами прекращения синтеза белка.
В процессе трансляции – синтезе белка из аминокислот, нуклеотидная последовательность м-РНК служит матрицей, с которой считываются триплеты, определяющие последовательность аминокислот. Синтез белка происходит при перемещении рибосомы по цепо чке м- РНК. Доставку аминокислот к комплексу рибосома – м-РНК вып олняют транспортные РНК (т-РНК). Для каждой аминокислоты имеется своя т- РНК, на одном конце которой находятся 3 неспаренных основа ния (антикодон), с помощью которого т-РНК выстраиваются в цепо чку, параллельную м-РНК, а к другому концу т-РНК присоединена аминокислота. В результате последовательного перемещения рибосомы по м-РНК растет синтезируемая цепь белка.
Все указанные процессы осуществляются при участии ферме нтов
– белковых катализаторов. Определяющими в консерватизме
Современный Гуманитарный Университет
25
наследственности являются точность воспроизведения мол екул ДНК при репликации, точность синтеза м-РНК при транскрипции и выс окая точность трансляции в синтезе белка.
Âпоследние десятилетия под геном понимали участок ДНК, кодирующий белковую цепочку или определяющий функциона льную молекулу РНК. В настоящее время различают структурные ген ы, которые кодируют белки или РНК, и регуляторные гены, которые регул ируют активность структурных генов, определяя их “включение” и “выключение”. В последнее время обнаружены участки повто ряющихся нуклеотидных последовательностей, функции которых мало изучены, обнаружены мигрирующие нуклеотидные последовательност и (мобильные гены). Все это вызывает ряд трудностей, связанн ых с определением границ гена в молекуле ДНК. Кроме того, струк тура гена имеет прерывистый характер. В нем выделяют экзоны – участки гена,
âкоторых закодирована информация для синтеза белка, они копируются в м-РНК, и интроны – участки, которые не содержат информации для синтеза белка, они участвуют в транскрипци и. Вначале ген копируется полностью в пре-м-РНК, а затем интроны вырез аются (процессинг), образуя зрелую м-РНК, которая используется в трансляции при синтезе белка, и соединяются (сплайсинг). Рассмотренны е выше процессы синтеза белка представлены на рис. 4.
Значительные успехи в области молекулярно-генетическог о исследования психики человека стали возможны благодаря появлению
â70-õ гг. такого экспериментального инструмента, как рестри кционные эндонуклеазы. Специальные ферменты обладают способност ью вступать
âреакцию с определенными участками (сайтами) в ДНК, котор ые называются сайты узнавания; и разрезать двухцепочечную м олекулу ДНК так, что одна из цепей ДНК оказывается на несколько нуклео тидов длиннее другой. Эти нуклеотиды, называемые также “липкими концами”, могут спариваться с комплементарными им нуклеотидами. Вс ледствие этого ДНК разных организмов могут объединяться, образуя т ак называемые рекомбинантные молекулы. Это свойство исполь зуют для размножения (амплификации) специфической, интересующей исследователя ДНК.
Âпрактическом аспекте важно то, что гены, контролирующие образование определенных белков, можно вводить в бактери и (клонировать гены) и быстро амплифицировать. Этот подход о снован на том, что в бактерии кроме своей кольцевой хромосомы, час то имеются дополнительные маленькие кольцевые молекулы двухцепоче чной ДНК, называемые плазмиды, которые воспроизводятся автономно . Плазмиды можно выделить и расщепить определенной рестриктазой та к, чтобы получить молекулу ДНК с “липкими концами”. Затем фрагмент ы ДНК человека с “липкими концами”, полученной после расщеплен ия такой же рестриктазой, можно сшить с плазмидной ДНК, используя д ля этого другой фермент – лигазу. Полученные таким образом плазмид ы вводят
âбактерии, где они размножаются.
Современный Гуманитарный Университет
26

Ядерная оболочка |
Плазматическая |
|
мембрана |
ßäðî
ÄÍÊ
Ядерные белки, |
|
связывающиеся |
|
ñ ÐÍÊ |
Сплайсинг РНК |
ТранспортРНК
Рибосомы |
Трансляция |
ì-ÐÍÊ
Белок
Цитоплазма
Рис.4. Синтез белка у эукариот (ДНК – РНК-белок). Благодаря ядерной оболочке активные рибосомы отделены от ядра; в результате транскрипты РНК проходят процессинг, до выхода из ядра в цитоплазму, где происходит трансляция. Таким образом, между транскрипцией ДНК и трансляцией РНК осуществляется процессинг и транспорт РНК
Современный Гуманитарный Университет
27
Первое положение |
|
Второе |
положение |
|
Третье по- |
|
|
|
|
|
ложение |
(5' конец) |
U |
C |
A |
G |
(3' конец) |
|
|
||||
U |
Phe |
Ser |
Tyr |
Cys |
U |
Phe |
Ser |
Tyr |
Cys |
C |
|
Leu |
Ser |
Cron |
Cron |
A |
|
Leu |
Ser |
Cron |
Trp |
G |
|
C |
Leu |
Pro |
His |
Arg |
U |
Leu |
Pro |
His |
Arg |
C |
|
Leu |
Pro |
Gln |
Arg |
A |
|
Leu |
Pro |
Gln |
Arg |
G |
|
A |
Ile |
Thr |
Asn |
Ser |
U |
Ile |
Thr |
Asn |
Ser |
C |
|
Ile |
Thr |
Lys |
Arg |
A |
|
Met |
Thr |
Lys |
Arg |
G |
|
G |
Val |
Ala |
Asp |
Gly |
U |
Val |
Ala |
Asp |
Gly |
C |
|
Val |
Ala |
Glu |
Gly |
A |
|
Val |
Ala |
Glu |
Gly |
G |
Рис. 5. Генетический код. При синтезе белка триплетынуклеотидовРНК (кодоны) транслируются в соответствующие им аминокислоты. Например,ок- äîíû CUG è CAG направляют в белок соответственновалин и глутаминовую кислоту. Обратите внимание, что кодоныU или С во втором положении обычно кодируют более гидрофобные аминокислоты
В настоящее время рестриктазы используются также для идентификации генов. Для этого разрезанные рестриктазой фрагменты ДНК идентифицируют с помощью библиотеки ДНК-зондов, котор ые представляют собой уникальные нуклеотидные последовате льности активно работающих генов или их частей. Часто для определ енной последовательности ДНК обнаруживается полиморфизм длин ы рестриктных фрагментов (ПДРФ), что является результатом р азличий в сайтах рестрикции у разных индивидов. В таких случаях ПДР Ф можно использовать для установления местоположения изучаемых генов в хромосомах при изучении сцепления генов в семьях. Количес тво локализованных генов в определенных районах хромосом че ловека при
Современный Гуманитарный Университет
28
анализе их сцепления с полиморфными участками ДНК постоя нно увеличивается. Технологические приемы молекулярной ген етики позволяют также определить последовательность нуклеоти дов в ДНК, то есть секвенировать ДНК. Для этого молекулу ДНК расщепляют с помощью рестриктазы на фрагменты. Затем определяют последовател ьность нуклеотидов во фрагментах и определяют с помощью специал ьных процедур очередность фрагментов в целой молекуле.
Таким образом, на основании данных о последовательности нуклеотидов и генетического кода можно определить последовательность аминокислот в полипептидной цепи (ри с.5), то есть определить белок, который контролирует данный ген. В псих огенетике, когда, как правило, неизвестны биохимические механизмы, вовлеченные в формирование психических свойств, техноло гии молекулярной генетики позволяют обнаружить такие гены и , следовательно, внести существенный вклад в раскрытие мех анизмов наследования психических свойств.
2.8. Виды изменчивости
Свойством, противоположным наследственности, является изменчивость, которая характеризует существование индивидов в разных формах и отражает различия этих форм. Различают на следуемые изменения самих генов (мутации), изменения, обусловленные сочетанием разных генов у индивидов (комбинативная наследственная изменчивость), изменения, вызванные влиянием средовых условий (модификационная изменчивость). Мутации могут быть обусловлены изменениями числа хромосом (утрата, лиш ние хромосомы) или изменениями части хромосом: делеция (утрата), дупликация (удвоение), инверсия (поворот участка на 180 град усов), транслокация (перенос участка одной хромосомы на другую) . Схема хромосомных изменений представлена на рис.6. Особый интер ес представляют точковые мутации, которые обусловлены изме нениями отдельных нуклеотидов. Точковые мутации представляют со бой вставки, выпадения, изменения нуклеотидов. В результате возникают новые аллели. Механизмы возникновения точковых мутаций предст авляют собой замену оснований или сдвиг рамки считывания.
Замена одного основания в цепи ДНК может привести к тому, ч то в синтезируемый белок будет включена другая аминокислота , и в результате функция белка может быть изменена. Проявление такой замены среди большого числа аминокислот зависит от струк туры и функции белка, местоположения аминокислоты.
Сдвиг рамки считывания связан с вставкой или выпадением о дного и более нуклеотидов. В результате случайного добавления и ли выпадения одного нуклеотида изменяется считывание (тран сляция) кодонов в м-РНК и, начиная с точки мутации, синтезируется из мененная последовательность аминокислот. Частота мутирования от дельных генов
Современный Гуманитарный Университет
29

в среднем равна 1/100000. Хотя частота новых мутаций мала, они являются материалом для естественного отбора в эволюцио нном процессе. Наследственность сохраняет не только сходство , но и мутационные различия индивидов.
A B C D
à
A B C
á
A B C D C D
â
A B D C
ã
E I Y C D
ä
Рис.6. Схема хромосомных изменений: а - нормальная (исходная) хромосома; б - делеция; в - дупликация; г - инверсия; д - транслокация
Комбинативная изменчивость обеспечивает многочисленно сть генотипов при половом размножении и определяет межиндив идуальные генетические различия. Даже для небольшого числа локусов , содержащих по несколько разных аллелей при рекомбинации вследствие перемешивания генных комплексов возникает м ножество генотипов. Если k – число локусов, m – число аллелей в каждом локусе, то возможное число генотипов при рекомбинации равно [(m(m+1)/2]k. Для 10 локусов, содержащих по 4 возможных аллеля, теоретичес ки
Современный Гуманитарный Университет
30