

Сжатая ZIP-папка / ЛК_ген_6
.docЛЕКЦИЯ 7 - ГЕНЕТИКА
ЛЕКЦИЯ 7
ГЕНЕТИКА ПОЛА
Исследование проблемы пола позволило вскрыть генетические механизмы определения и становления одного из сложных признаков организма в онтогенезе. При этом, как оказалось, решение этого вопроса имело не только практический, но и значительный теоретический интерес, позволивший достаточно глубоко проникнуть в понимание процессов наследственности и изменчивости.
Половые признаки делят на первичные и вторичные. К первичным относят морфологические и физиологические особенности организма, которые обеспечивают образование гамет и их соединение в процессе оплодотворения. Это – гонады, половые пути и наружные гениталии у высших животных, андроцей и гинецей – у высших растений.
Вторичные половые признаки и свойства организма непосредственно не обеспечивают процессы гаметогенеза, спаривания и оплодотворения, но играют вспомогательную роль в половом размножении. Это – особенности строения плавников у рыб, грудных желёз у млекопитающих, сроки цветения у высших растений и т.д.
Половой диморфизм широко распространён в природе и встречается на всех ступенях эволюционного развития. Его биологическая роль состоит в перекомбинации наследственной информации, что ведёт к увеличению общего фонда наследственной изменчивости.
ХРОМОСОМНЫЙ МЕХАНИЗМ ОПРЕДЕЛЕНИЯ ПОЛА
Известно, что соотношение полов у раздельнополых организмов близко к отношению 1:1. Поэтому было высказано предположение о том, что один из полов должен быть гомозиготным, а другой – гетерозиготным. Первое экспериментальное доказательство этой гипотезе было получено известным учёным К.Корренсом. В конечном итоге, пол, образующий одинаковые в отношении определения пола гаметы, назвали гомогаметным, а пол, образующий разные гаметы – гетерогаметным.
Цитологические исследования показали, что у большинства животных и раздельнополых растений в хромосомном наборе мужских и женских особей хромосомы одной из пар значительно отличаются друг от друга, или одна из хромосом представлена в единственном числе.
Позднее было установлено, что с этими необычными хромосомами связано определение пола, поэтому их стали называть половыми. Обычные хромосомы называют аутосомами, а половые представлены хромосомами, обозначаемые буквами Х и У.
Известны 4 основных типа хромосомного определения пола.
|
Тип определения |
Гетерогамет-ный пол |
Гаметы |
Зиготы |
Организмы |
||
|
Мужской |
Женский |
Мужской |
Женский |
|||
|
ХУ |
Мужской |
Х и У |
Х |
ХУ |
ХХ |
Человек, млекопитающие, дрозофила и др. |
|
ХО |
Мужской |
Х и О |
Х |
ХО |
ХХ |
Кузнечики, клопы |
|
ХУ |
Женский |
Х |
Х и У |
ХХ |
ХУ |
Птицы, земноводные, рептилии, бабочки, клубника и др. |
|
ХО |
Женский |
Х |
Х и О |
ХХ |
ХО |
Моль и др. |
БАЛАНСОВЫЙ МЕХАНИЗМ ОПРЕДЕЛЕНИЯ ПОЛА
По мере накопления фактов, хромосомная теория определения пола не всегда находила такие строгие подтверждения. Напр., иногда встречались гинандроморфы – особи, совмещавшие в себе признаки как мужского, так и женского организмов.
В 20-е годы ХХ столетия американский исследователь К.Бриждес обнаружил, что развитие признаков пола у плодовой мушки дрозофилы сильно изменяется в зависимости от соотношения Х-хромосом и аутосом в генотипе особи. Среди таких мушек иногда возникали самки с триплоидным набором хромосом: три набора аутосом (А) и три Х-хромосомы (3А+3Х). Некоторые из таких триплоидных самок были плодовиты, но, как можно предположить, в мейозе у них нарушено расхождение хромосом. В силу этой причины при скрещивании таких самок с диплоидными самцами в потомстве было получено 8 типов особей, имевших различное соотношение половых хромосом и аутосом, и соответствующие им половые признаки.
На основании таких экспериментов Бриджес пришёл к выводу о том, что у дрозофилы женский пол определяется вовсе не наличием двух Х хромосом, а развитие мужского пола – не сочетанием Х и У хромосом, как записано в таблице. Половые признаки мушки в таких случаях определялись отношением числа Х хромосом к числу наборов аутосом, т.е. Х:А.
При отношении Х:А < 0,5 из зиготы развиваются так называемые сверхсамцы,
при отношении Х:А = 0,5 – нормальные самцы,
при 0,5 < X:A < 1 – интерсексы,
при Х:А = 1 – нормальные самки и
при X:A > 1 – сверхсамки.
Полученные результаты легли в основу балансовой теории определения пола у организмов. Вполне естественно, что сверхсамки и сверсамцы, обладавшие внешними признаками соответствующего пола, а также интерсексы, оказывались бесплодными вследствие глубоких нарушений процесса гаметогенеза у таких особей.
Многочисленные экспериментальные результаты позволили подтвердить эту теорию, а также придти к выводу о том, что гены, определяющие половые признаки организма, находятся в аутосомах, а гены, обусловливающие функционирование аутосомных генов пола, могут находиться как в половых хромосомах, так и в аутосомах. Т.е. половым хромосомам отведена регуляторная роль в формировании половых признаков особи.
У человека нерасхождение половых хромосом при образовании яйцеклеток и образование зиготы при слиянии такой яйцеклетки с нормальным сперматозоидом вызывает у сформировавшейся особи тяжёлые физические и психические заболевания. Известны следующие хромосомные болезни человека этого типа:
44 А + ХХХ – трисомия по Х хромосоме,
44 А + ХХУ – синдром Кляйнфельтера,
44 А + Х – синдром Шерешевского – Тернера,
44 А + У – нежизнеспособные зиготы, отмирающие в самом начале развития зародыша.
Все остальные особи часто не доживают до половозрелости и не имеют потомков.
РОЛЬ УСЛОВИЙ СРЕДЫ В ОПРЕДЕЛЕНИИ ПОЛА
В природе выделяют три группы организмов.
У первой группы, к которой относится большинство известных раздельнополых организмов, условия среды не контролируют пол развивающейся особи. Он определяется только генетическим механизмом.
Вторая группа, к которой относят некоторых морских беспозвоночных, характеризуется механизмом определения пола, обусловленного условиями среды. В таких случаях индифферентные в половом отношении личинки червя могут прикрепиться к хоботку самки, и в таком случае они превращаются в самцов. Если такие личинки ведут свободный образ жизни, в конечном итоге они прикрепляются к субстрату и превращаются в самок.
Третья группа организмов включает обоеполые растения и гермафродитных животных, у которых сохранился этот наиболее древний тип определения пола. Он осуществляется в ходе онтогенетической дифференцировки наследственно идентичной ткани в сторону либо мужского, либо женского пола. Зачаточные индифферентные в половом отношении гонады у эмбрионов таких животных имеют двойственную природу и состоят из внутренних и внешних слоёв. В ходе дифференциации пола идёт развитие одного из слоёв гонады и подавление развития другого. Этот процесс регулируется гормонами, выделяемыми различными железами, а уровень гормональной секреции контролируется генами. Функционирование таких генов часто зависит от условий внешней среды. Экспериментально показано, что гормоны у таких организмов могут приводить к формированию самок с соответствующими органами размножения из генотипических самцов, и наоборот.
ПРАКТИЧЕСКИЕ ВОЗМОЖНОСТИ УПРАВЛЕНИЯ ПОЛОМ
Особенности различных отраслей сельскохозяйственного производства делают необходимым предпочтительное разведение особей мужского, либо женского пола. Напр., в птицеводстве важно разведение кур, в молочном производстве – телочек, а в мясном скотоводстве – бычков и т.п. В этом аспекте генетика напрямую оказывается связанной с экономикой и повышением эффективности производства, что делает весьма актуальной разработку приёмов и методов распознавание особей по признакам пола на самых ранних этапах онтогенеза. Из имеющихся положительных примеров в этой сфере можно отметить следующие.
У тутового шелкопряда путём генетических скрещиваний удалось перенести ген окраски грены (яиц) в генотип самок. Благодаря этому уже на самой ранней стадии удаётся проводить отбор будущих самцов, которые образуют коконы, содержащие на 25-30% натурального шёлка больше, чем коконы, образуемые самками.
Для крупного рогатого скота показано, что при проведении электрофореза спермы сперматозоиды распределяются в среде под действием электрического поля таким образом, что в зоне с рН около 6,8 скапливаются, в основном, сперматозоиды, несущие Х-хромосому, а в области с рН около 7,7 – сперматозоиды с У-хромосомой. Искусственное осеменение, используемое в современном сельскохозяйственном производстве, значительно повышает вероятность рождения особей ожидаемого пола.
НАСЛЕДОВАНИЕ ПРИЗНАКОВ, СЦЕПЛЕННЫХ С ПОЛОМ
Это направление генетики развивалось одним из первых, и именно результаты в этой области исследований дали множество различных доказательств того, что носителями генетической информации являются хромосомы, а затем и – гены. Исследование особенностей мейоза и знания о том, что в разные гаметы попадают разные половые хромосомы, сделало возможным проследить за наследованием признаков, обусловленных присутствием половых хромосом. Поэтому далее стали говорить о наследовании, сцепленном с полом.
Такое явление было выявлено и подробно изучено в лаборатории Т.Моргана. Напр., при изучении окраски глаз у дрозофилы было установлено, что красный цвет (W) доминирует над белым (w). При скрещивании красноглазых самок с белоглазыми самцами в первом поколении всё потомство оказывалось красноглазым.
При скрещивании особей первого поколения в F1 все самки оказались красноглазыми, а среди самцов – половина имела красную окраску глаз, а половина – белую.
При скрещивании белоглазых самок с красноглазыми самцами в первом поколении все самки оказались красноглазыми, а самцы, наоборот, - белоглазыми. В F2 – половина самок и ½ самцов имели белые глаза, а остальные – красные.
Полученные закономерности становятся понятными, если предположить, что гены, определяющие окраску глаз у дрозофилы, находятся только в половых Х-хромосомах, а в Y-хромосомах – их нет. В таком случае запись выявленных закономерностей имеет следующий вид:
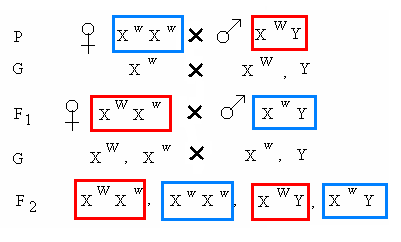
Таким образом, гены окраски глаз и гены, определяющие развитие признаков женского пола, находятся в одной и той же хромосоме и поэтому передаются сцепленно.
Наследование признаков, сцепленных с полом, при гетерогаметности женского пола можно проанализировать таким же образом. Напр., у кур полосатое оперение определяется доминантным геном В. Решётки Пеннета будут аналогичны приведённым выше и отличаться только тем, что куры имеют генотип XY, а петухи – ХХ.
Изучение наследования рецессивного признака жёлтой окраски тела у мухи дрозофилы методом генетического анализа так же привело к выводу о том, что этот признак связан с половыми хромосомами.
Таким образом, экспериментально было доказано явление сцепленного с полом наследования ряда признаков у разных организмов.
Касаясь человека, следует сказать о том, что в связи с Х-хромосомой у него наследуется около 60 генов. Среди них встречаются как доминантные, так и рецессивные по проявлению признаки. К доминантным относят, напр., рахит, не поддающийся лечению витамином D, тёмную эмаль зубов и т.д. Среди рецессивных можно отметить гемофилию, дальтонизм (неспособность различать красный и зелёный цвета), атрофию зрительного нерва, миопатию Дюшена. При этом указанные рецессивные признаки проявляются лишь у мужчин, в силу того, что Y-хромосома не имеет аллелей этих признаков. Напр., распишем наследование гемофилии, при котором рецессивный аллель содержится в одной Х-хромосоме.
Рисунок.
Принятые обозначения указывают на то, что женщина с генотипом ХhХh также страдает гемофилией. Действительно описаны семьи, в родословных которых имеются близкородственные браки, что и привело к появлению гемофилии у женщин. При этом признак у них выражен примерно вдвое слабее, чем у мужчин с той же болезнью, что можно объяснить взаимодействием с какими-либо иными генами женского организма.
Поэтому в медицинской литературе не всегда можно встретить примеры достаточно чёткого проявления у лиц разного пола тех или иных синдромов, связанных с половой Х-хромосомой.
Ряд признаков сцеплен с половой Y-хромосомой. Это гены локализованы в тех районах Y-хромосомы, которые не имеют аналогов в Х-хромосоме. Поэтому они определяются только одним аллелем, присутствующим в организме мужчины, и передаются от отца к сыну. Среди таких голандрических признаков отмечают гипертрихоз – наличие волос по краю ушной раковины, синдактилию, проявляющуюся в виде кожистых перепонок между пальцами ног и т.д. Изображение схемы наследования таких признаков не составит значительного труда.
В то же время гомологичные участки половых Х и Y хромосом содержат ряд аллельных генов, которые проявляются у представителей обоих полов. Ярким примером такого признака является общая цветовая слепота (отсутствие цветового зрения). Несмотря на его рецессивный характер, он проявляется как у мужчин, так и у женщин. Поэтому такие признаки наследуются по классическим законам Менделя.
Однако, встречаются и признаки, сцепленные с половыми хромосомами, но проявление которых ограничено половой принадлежностью особи, либо регулируется ею.
К первой группе признаков можно отнести, напр., гены, определяющие ширину таза, которые наследуются и от отца, и от матери, но проявляются в полной мере только у детей женского пола. Среди мужских признаков, ограниченных полом, является количество и распределение волосяного покрова на теле, которое наиболее ярко проявляется у сыновей.
Другая категория признаков, контролируемая или зависимая от пола, по-разному проявляется у мужчин и женщин. Напр., раннее облысение у мужчин проявляется как доминантный признак, поэтому как гомозиготы – АА, так и гетерозиготы – Аа имеют одинаковый фенотип. И их относительно больше, чем гомозигот – аа. У женщин этот признак проявляется как рецессивный. Поскольку относительное число гомозигот – аа – значительно меньше, то и отмеченный фенотип встречается у женщин не столь часто. Кроме того, свои коррективы в проявление этого признака у женщин вносит, по-видимому, и взаимодействие с другими генами, что в целом значительно снижает число особей женского пола с отмеченным нарушением.
Ещё одним примером зависимости проявления признака от пола у человека является болезнь – подагра, пенетрантность которой у мужчин составляет около 80%, в то время как у женщин – только 12%. Следовательно, болезнь чаще проявляется у мужчин.
Степень выраженности признаков – экспрессивность также может быть неодинаковой. Напр., тембр голоса не только отличается у разных представителей одного пола, но и меняется в ходе онтогенеза. Развитие таких признаков связывают с половыми гормонами, уровень которых меняется как в процессе развития организма, так и различен у разных особей.
Таким образом, наследование признаков, сцепленных с полом у человека, не всегда однозначно и может зависеть от ряда факторов.