

MolBiol_sivolob
.pdfСиволоб А.В. Молекулярна біологія
тивності. Альтернативою авторадіографії є експонування гелю на спеціальний фосфорний екран, який потім сканується сполученим із комп'ютером приладом – фосфоріміджером. Крім вищої, якщо порівнювати з авторадіографією, чутливості, використання фосфоріміджера має ту перевагу, що дозволяє зі значно вищою точністю оцінювати кількість матеріалу в смузі.

 Фрагменти ДНК
Фрагменти ДНК
Полімерна сітка геля
-
+
Рис. 11.4. Розділення фрагментів ДНК за розміром шляхом гель-електрофорезу. Праворуч – окремі зони, що містять розділені фрагменти
У найпростішому варіанті – для розділення фрагментів дволанцюгової ДНК за розміром – найчастіше використовують агарозні гелі. Поліакриламідний гель (ПААГ) має значно різноманітніше застосування. Зокрема, саме ПААГ використовують для розділення сумішей білків. Найпопулярнішим є варіант електрофорезу білків у присутності додецилсульфату натрію – зарядженого детергента, який розгортає поліпептидний ланцюг і вкриває його своєрідною “шубою”. У результаті відбувається розділення ланцюгів різної довжини за принципом, що ілюструє рис. 11.4.
На рухливість макромолекул під час електрофорезу впливає також їхня форма. Відповідно, за допомогою електрофорезу можна, зокрема, розділити:
•Молекули ДНК, що містять перманентні вигини, зумовлені особливою послідовністю пар основ – вигнута ДНК більшою мірою гальмується полімерною сіткою гелю.
336

Розділ 11. Методи молекулярної біології
•Циркулярні та лінійні молекули ДНК, а також окремі циркулярні топоізомери (див. розділ 3), оскільки вони розрізняються за ступенем компактності.
•Молекулу вільної ДНК і комплекс цієї ДНК із білком – зрозумі-
ло, що комплекс більше гальмується сіткою гелю.
Особливим варіантом електрофорезу нуклеїнових кислот є електрофорез одноланцюгових фрагментів ДНК у денатуруючих умовах (підвищена температура, висока концентрація сечовини). Якщо такий гель (поліакриламідний) досить тонкий (~0,4 мм – секвенуючий гель), він дозволяє розділити фрагменти, довжина яких різниться на один нуклеотид.
Створення та скринінг геномних бібліотек
Раніше було розглянуто принципи клонування фрагмента ДНК. Але звідки береться цей фрагмент? Для того, щоб працювати з певною ділянкою ДНК (наприклад, певним геном) спочатку здійснюють клонування великої кількості різноманітних фрагментів геному – створюють бібліотеку клонів, серед яких уже шукають потрібний.
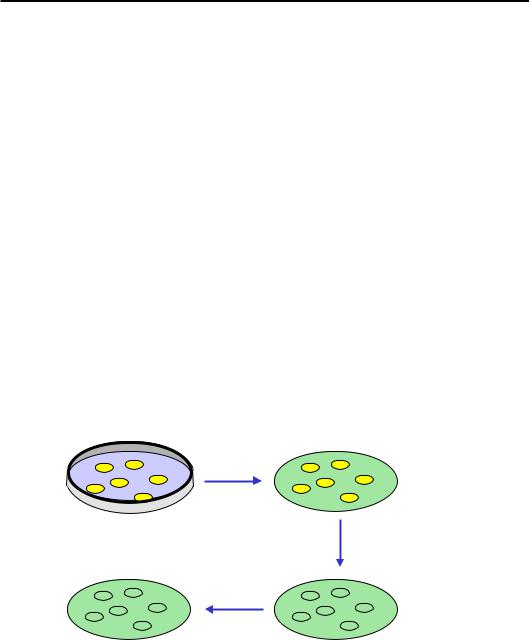
Стандартна процедура створення бібліотеки – метод дробовика (shotgun) – розпочинається з часткової (протягом обмеженого часу) обробки всієї геномної ДНК певною рестриктазою – таким чином, щоб отримати набір різноманітних фрагментів, які перекриваються, середня довжина яких становить ~20 тис. пар основ (рис. 11.5). Усі такі фрагменти вбудовують у вектори, і здійснюють трансформацію. Бактерії висівають на тверде середовище так, що кожна колонія походить від однієї клітини. Отже, набір колоній – це набір різноманітних клонів, кожен з яких містить один із фрагментів геному.
Геномна ДНК |
|
|
|
|
|
|
|
Рестриктні сайти |
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Продукти
часткової
деградації
Рис. 11.5. Метод дробовика
337
Сиволоб А.В. Молекулярна біологія
Аналогічно створюють бібліотеки клонів кДНК (комплементарна ДНК-копія молекули мРНК). Для цього спочатку виділяють сумарну мРНК із клітин певного типу. Використовуючи зворотну транскриптазу та оліго(dT), які є комплементарними 3'-кінцевим polyA-хвостам мРНК, як праймери, на цих мРНК синтезують комплементарні ланцюги ДНК. Видаляють ланцюг РНК РНКазою і добудовують другий ланцюг ДНК за допомогою фрагмента Кленова ДНК-полімерази І. Далі отримані молекули кДНК піддають клонуванню. На відміну від геномної бібліотеки, бібліотека клонів кДНК містить тільки кодуючі послідовності (екзони) генів і тільки таких, котрі є активними в клітинах даного типу.
Потрібну нуклеотидну послідовність серед бібліотеки клонів можна знайти за допомогою гібридизації клонованої ДНК із радіоактивноміченим ДНК-зондом (рис. 11.6). Колонії бактерій у чашці Петрі переносять на нітроцелюлозний фільтр – роблять своєрідну репліку. Здійснюється лізис клітин, депротеїнізація та денатурація ДНК у лужному розчині. Після висушування фільтра одноланцюгова ДНК необоротно фіксується на ньому. Фільтр обробляють радіоактивно-міченою ДНК певної послідовності – зондом. Якщо зонд є комплементарним ДНК клону, відбувається гібридизація – ренатурація дволанцюгової ДНК. Детектування цього факту за допомогою авторадіографії дозволяє ідентифікувати розшукуваний клон.
Перенесення колоній на нітроцелюлозний фільтр
Депротеїнізація, денатурація ДНК
Обробка зондом
Рис. 11.6. Гібридизація клонованої ДНК із радіоактивним зондом на нітроцелюлозному фільтрі
338

Розділ 11. Методи молекулярної біології
Зрозуміло, що описаний скринінг бібліотеки клонів залежить від наявності зонда. Одна з можливостей приготувати зонд базується на відомостях про амінокислотну послідовність білка того гена, клон якого потрібно відшукати (або білка-гомолога іншого організму). Хімічний синтез короткого олігонуклеотиду (~20 нуклеотидів) із послідовністю, що відповідає фрагменту амінокислотної послідовності білка й тому має бути комплементарною ділянці ДНК клону, дозволяє отримати зонд. Після внесення радіоактивної мітки такий олігонуклеотид можна використати для гібридизації. Проте, зважаючи на виродженість генетичного коду, бажано синтезувати набір послідовностей, які відповідають усім можливим комбінаціям кодонів для даної ділянки послідовності амінокислот. Суміш таких альтернативних олігонуклеотидів, один із яких є напевно комплементарним, і використовують як зонд.
Замість синтезу набору олігонуклеотидів можна обійтися синтезом лише одного зонда, використовуючи базу даних EST (Expressed Sequence Tag). База є загальнодоступною колекцією великої кількості коротких (200–400 пар основ) фрагментів послідовностей кДНК багатьох організмів. Щоб приготувати зонд, достатньо знайти в EST-базі коротку ділянку, яка відповідає фрагменту послідовності білка, і синтезувати її.
Полімеразна ланцюгова реакція
Альтернативним і додатковим до клонування методом збільшення кількості бажаного фрагмента ДНК – ампліфікації – є полімеразна ланцюгова реакція (ПЛР, або PCR – Polymerase Chain Reaction). Практично ПЛР – це реакція синтезу ДНК in vitro, яка повторюється багато разів: синтезовані ланцюги стають матрицями для синтезу в наступному циклі реакції. Для здійснення ПЛР треба знати принаймні короткі послідовності ДНК на кінцях того фрагмента, котрий має бути ампліфікований.
Реакційна суміш містить слідові кількість вихідної ДНК, надлишкові кількості двох синтетичних праймерів до кінців фрагмента, нуклеозидтрифосфати чотирьох типів і ДНК-полімеразу Taq термофільної бактерії Thermus aquaticus. Використання саме полімерази Taq зумовлено тим, що вона не втрачає своєї активності при нагріванні до високих температур.
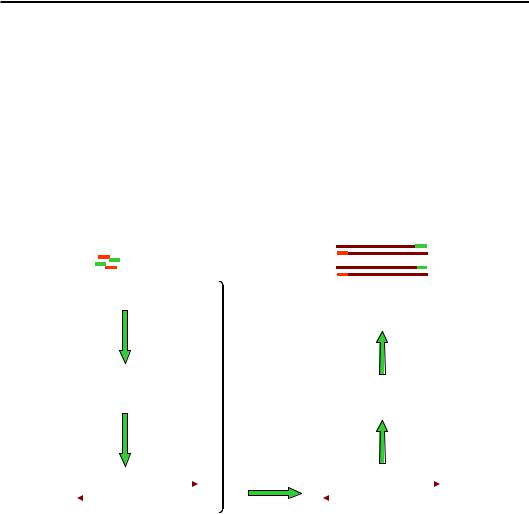
У першому циклі ПЛР (рис. 11.7) відбувається нагрівання суміші до 95 °С (денатурація ДНК) і охолодження до 60 °С. Після охолодження до такої все ще досить високої температури ланцюги ДНК не ренатурують, але праймери (оскільки вони присутні в досить високих кон-
339
Сиволоб А.В. Молекулярна біологія
центраціях) знаходять свої комплементарні ділянки й зв'язуються з ними. Полімераза Taq починає працювати, подовжуючи ці праймери.
Усі наступні цикли ПЛР (реакція здійснюється в автоматичному режимі на приладі, що називається ампліфікатором) полягають у нагріванні суміші до 95 °С, яке змінюється охолодженням до 60 °С. Після другого циклу будуть знов синтезовані дві такі самі молекули, як після першого, а також дві молекули, котрі зображено на рис. 11.7. Після третього циклу – дві такі самі молекули, що й після другого, а також дві молекули, довжина яких чітко обмежена двома праймерами (рис. 11.7). Далі кількість таких коротких матриць буде зростати, і після 30-го циклу їх буде в мільйон разів більше, ніж вихідних довгих матриць.
Праймери
ДНК
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Нагрівання |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
||||
|
|
до 95°С, |
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
||||
|
|
охолодження |
|
|
|
4 цикл |
||||||
|
|
до 60°С |
|
|
|
|||||||
|
|
|
|
|
|
1 цикл |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Синтез ДНК |
|
|
|
|
|
|
|
|||
|
|
|
|
|
3 цикл |
|||||||
|
|
при 60°С |
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
2 цикл |
|
|
|
|||
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Рис. 11.7. Полімеразна ланцюгова реакція. Деякі матриці, довжина яких виходить за межі ділянки,
обмеженої двома праймерами, не показано (кількість таких матриць після багатьох циклів залишається дуже малою)
Метод ПЛР має надзвичайно широке застосування. Він використовується кожного разу, коли є необхідність детектувати та дослідити невелику кількість ДНК, у тому числі – у складі неочищеного біологічного матеріалу. ПЛР застосовують також у комбінації з клонуванням: ампліфікований ДНК-продукт можна клонувати й використати, наприклад,
340

Розділ 11. Методи молекулярної біології
для експресії білка (див. нижче); ампліфікація за допомогою ПЛР стає в нагоді для збільшення невеликої кількості клонованої ДНК.
ПЛР застосовують також для ампліфікації мРНК у вигляді її кДНК-копій – після синтезу кДНК на мРНК за допомогою зворотної транскриптази: варіант ПЛР, що позначається як RT-PCR (Reverse Transcriptase Polymerase Chain Reaction). Абревіатура RT-PCR вико-
ристовується також у іншому значенні – Real Time Polymerase Chain Reaction (інше позначення: QRT-PCR – Quantitative Real Time Polymerase Chain Reaction): процедура, в якій ампліфікація ДНК супроводжується визначенням її кількості після кожного раунду за допомогою флуоресцентних барвників.
Секвенування ДНК
Клонований або ампліфікований фрагмент ДНК можна дослідити різними способами, однак найвичерпнішу інформацію дає встановлення нуклеотидної послідовності (sequence) фрагмента – секвенування.
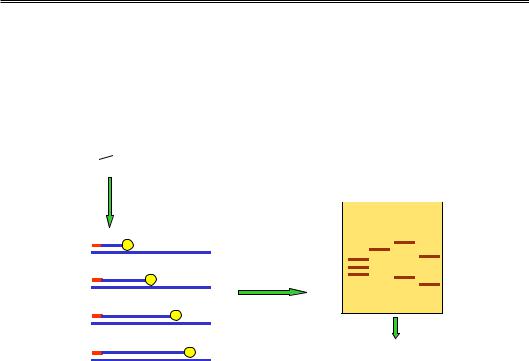
На рис. 11.8 показано схему найбільш популярного сьогодні методу Сангера (Frederick Sanger). До одноланцюгової ДНК-матриці додається радіоактивно мічений праймер, повний набір дезоксинуклеозидтрифосфатів (dNTP), ДНК-полімераза та невелика кількість дидезоксинуклеозидтрифосфату одного із чотирьох типів (наприклад, ddATP). Дидезоксинуклеотид відрізняється тим, що містить атом Н замість ОН-групи не тільки при 2'-, а також і при 3'-атомі пентози (див. рис. 3.2). Включення такого нуклеотиду в ланцюг, що синтезується, приведе до зупинки подальшого зростання ланцюга внаслідок відсутності 3' ОН-групи на його кінці (див. рис. 3.4). Оскільки ddATP присутній у невеликій кількості, така подія буде відбуватися в різних точках ланцюга – в усіх, де аденін розташований напроти тиміну в складі матриці. Шляхом денатурації продуктів реакції отримаємо набір мічених одноланцюгових фрагментів від праймера до кінцевого аденіну. Довжина цих фрагментів у нуклеотидах визначить порядковий номер аденіну в складі ланцюга.
З метою визначення довжини фрагментів проводять гель-електро- форез у денатуруючих умовах, на сусідні лунки гелю наносять також продукти синтезу у присутності інших дидезоксинуклеотидів. Як показано на рис. 11.8, після електрофорезу та візуалізації смуг із такого гелю можна прочитати нуклеотидну послідовність.
341
Сиволоб А.В. Молекулярна біологія
В іншому методі секвенування – методі Максама – Гілберта (Allan M. Maxam, Walter Gilbert) замість ферментативного синтезу застосовують хімічне розрізання ланцюга на нуклеотиді певного типу. Отримані фрагменти різної довжини так само розділяють за допомогою електрофорезу.
праймер
5'
|
|
|
|
|
|
|
|
|
ДНК-полімераза + |
A G T C |
|||||
|
dATP, dTTP, dCTP, dGTP + |
||||||
|
невелика кількість ddATP |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
А
А
АДенатурація, гель-електрофорез
А |
CTAAACGTGA |
Рис. 11.8. Секвенування ДНК за Сангером: схема синтезу ДНК у присутності дидезоксиАТР (ліворуч). Шляхом аналогічної процедури для інших трьох дидезоксинуклеотидів отримуємо набір одноланцюгових фрагментів,
що аналізуються за допомогою гель-електрофорезу в денатуруючих умовах (праворуч) – розподіл смуг дозволяє прочитати послідовність (праворуч унизу)
Метод Сангера використовується сьогодні автоматичними секвенаторами, в яких замість радіоактивно-мічених застосовуються флу- оресцентно-мічені праймери. Для кожної з чотирьох реакцій беруть чотири різні флуоресцентні мітки, котрі випромінюють світло в різних спектральних діапазонах. Продукти всіх чотирьох реакцій наносять на гель для електрофорезу разом. Сканування гелю після електрофорезу лазерним променем, що збуджує флуоресценцію, дозволяє дискримінувати продукти різних реакцій, тобто різні кінцеві нуклеотиди, і, таким чином, негайно прочитати послідовність.
Здійснення секвенування кожного із фрагментів, які містяться в геномній бібліотеці клонів, і порівняння послідовностей фрагментів, що перекриваються (рис. 11.5), дозволяє розмістити клоновані фрагменти в порядку їхньої локалізації в геномі, тобто встановити повну послідовність геному.
342

Розділ 11. Методи молекулярної біології
Інший сучасний підхід у секвенуванні геномів (так зване піросеквенування), який реалізується на автоматизованих секвенаторах, дозволяє визначити послідовність значно швидше, дешевше і при цьому не потребує ані клонування ДНК, ані електрофорезу. Одноланцюгові фрагменти, отримані з невеликої кількості геномної ДНК, пришивають 5'-кінцями до мікрокульок (один фрагмент на кульку) та ампліфікують за допомогою ПЛР. Кожну кульку із пришитими до неї ампліфікованими ідентичними фрагментами розміщують у мікрореакторі, де здійснюється ДНК-полімеразна реакція. Нуклеозидтрифосфати подаються в реакційну суміш імпульсно один за одним. Якщо нуклеотид певного типу виявляється комплементарним матриці та включається у зростаючий ланцюг, пірофосфат, що при цьому звільняється, залучається до низки хімічних реакцій, де остання реакція супроводжується випромінюванням світла (хемілюмінесценція). Світловий сигнал фіксується оптичною системою, і послідовність таких сигналів читається як нуклеотидна послідовність. Реакція здійснюється паралельно у 200 тис. мікрореакторів (для 200 тис. фрагментів, які перекриваються), що дозволяє встановити послідовність приблизно 200 млн пар основ за 4,5 години.
Останнім часом з'явилася нова перспектива секвенування ДНК у процесі протягування молекули через нанопору. Пóра діаметром ~1 нм утворюється у кремнієвій мембрані, і одноланцюгова ДНК досить повільно проходить крізь цю пору під дією електричного поля. Виявилося, що характеристики електричного струму через мембрану залежать від того, який нуклеотид знаходиться в порі в даний момент. Є сподівання, що з розвитком цього методу з'явиться можливість швидкого та дешевого секвенування геному однієї конкретної людини.
Біоінформатика
Велика кількість послідовностей (у тому числі повних послідовностей геномів), які вже встановлені та продовжують накопичуватись, створює проблему їхнього збереження та аналізу. Зрозуміло, що таке питання, яке постає вже при порівнянні послідовностей окремих клонованих фрагментів з метою з'ясування нуклеотидної послідовності геному, може бути вирішеним лише за допомогою комп'ютерів. Біоінформатика – галузь, пов'язана з розв'язанням подібних проблем, є сьогодні невід'ємною складовою частиною молекулярної біології.
Найбільші загальнодоступні через Інтернет бази даних нуклеотидних і амінокислотних послідовностей створено в Європейській Лабораторії Молекулярної Біології (EMBL Sequence Data Base) і Національних Інститутах Здоров'я США (GenBank). На відповідних порталах розміщено
343

Сиволоб А.В. Молекулярна біологія
також програмні засоби порівняння послідовностей, пошуку промоторів, стартових точок транскрипції, інтронів та екзонів, визначення білкових послідовностей із послідовностей нуклеотидів тощо. Серед баз даних on-line слід згадати також бази даних просторових структур білків
(Protein Data Bank) і нуклеїнових кислот (Nucleic Acid Database).
На основі біоінформатичного аналізу можна встановити, наприклад, функціональне значення невідомого щойно клонованого гена за його гомологією з відомим геном іншого організму, структуру гена (знайти промотор, екзони, точки термінації транскрипції), структурнофункціональні особливості нових білків, функціональні зв'язки між різними білками та генами, філогенетичні стосунки між різними таксонами. Завдяки розвитку біоінформатики існує можливість (у доповнення до результатів, що отримують in vivo та in vitro) розв'язувати різноманітні проблеми просто за допомогою комп'ютера – in silico.
Експресія рекомбінантних білків
Оскільки переважна більшість білків синтезується клітиною в невеликих кількостях, просте виділення цих білків із метою їхнього дослідження є практично неможливим. Технології рекомбінантних ДНК вирішують це питання. Ген, що кодує даний білок, можна клонувати, вбудувати (наприклад, у бактеріальну плазміду) і змусити бактеріальну культуру продукувати цей білок. Виділення та очищення білка з культури біохімічними методами вже не є принциповою проблемою, зважаючи на його велику кількість.
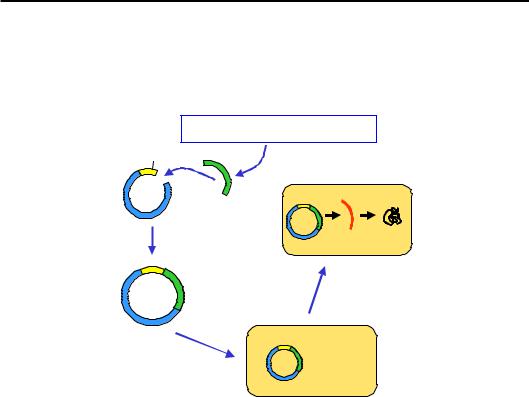
Одну з найпростіших схем експресії білка в E. сoli зображено на рис. 11.9. Зрозуміло, що еукаріотичний ген не має сенсу вводити в прокаріотичну клітину – прокаріоти не мають системи сплайсингу. Тому беруть лише кодуючу частину гена, яку можна отримати з бібліотеки клонів кДНК. Використовуючи придатну рестриктазу та лігазу, потрібну кДНК вбудовують у плазміду поряд із промотором – наприклад, лактозного оперона (див. розділ 5). Здійснюють трансформацію рекомбінантної плазміди в бактеріальні клітини, до бактеріальної культури додають синтетичний індуктор lac-оперона IPTG. Промотор активується, і здійснюється транскрипція гена та трансляція білка.
Більш ефективна двостадійна система експресії використовує промотор РНК-полімерази бактеріофага Т7. У бактеріальному геномі ні таких полімераз, ні відповідних промоторів немає. У клітину вводяться два плазмідні вектори: один містить lac-промотор і ген РНК-полі-
344
Розділ 11. Методи молекулярної біології
мерази Т7, інший – сильний промотор полімерази Т7 разом із геном білка, що має бути експресований. IPTG індукує експресію полімерази, яка зв'язується тільки з промотором у складі другої плазміди (інших промоторів немає) і забезпечує синтез великої кількості білка.
Бібліотека клонів кДНК
lac промотор
мРНК білок
Плазмідний
вектор
+ IPTG
Трансформація
Рис. 11.9. Експресія білка в бактеріальній клітині. lac-Промотор – промотор лактозного оперона, IPTG – синтетичний індуктор оперона
Вивчення структури й особливостей функціонування багатьох білків було б просто неможливим без такої штучної експресії. Крім того, використання рекомбінантних ДНК для експресії білків відкриває принципово нові можливості: послідовність ДНК можна практично як завгодно змінити й отримати мутантну форму білка. До білкового гена можна пришити ген іншого білка, який буде виконувати роль мітки. Частину гена, що відповідає частині білка (скажімо, структурному домену) можна вилучити, й отримати редукований білок чи окремий домен для структурних досліджень. Будь-який нуклеотид можна замінити на інший методами спрямованого мутагенезу, завдяки чому отримати білок з амінокислотною заміною. Такі підходи мають велике значення для вивчення механізмів активності ферментів, специфічного впізнання білками інших молекул тощо.
345