

Глава 2 слуховая сенсорная мистема
В связи с возникновением речи как средства межличностного общения, слух у человека играет особую роль, поскольку именно его слуховая система предназначена преимущественно для того, чтобы слышать речь других людей. Потерю слуха нельзя недооценить. Известно, что глухота или даже лишь недостаток слуха представляют собой наиболее серьезную угрозу для умственного развития ребенка, так как именно словесное общение является важнейшим средством обучения. Слух требуется и для умения говорить самому, поэтому глухие от рождения дети не научаются говорить, так как лишены слуховых раздражителей. По этой причине глухота, возникающая до приобретения речи, относится к особенно серьезным проблемам, и тугоухие от рождения дети должны начинать пользоваться слуховым аппаратом до 1,5 лет.
►Морфология и физиология периферического отдела слухового анализатора. Периферическая часть слухового анализатора анатомически тесно связана с периферической частью вестибулярного анализатора – они располагаются рядом в каменистой части (пирамиде) височной кости (внутреннее ухо). Кроме того, они имеют и филогенетическое «родство» - происходят из одних и тех же структур в процессе эволюции.
Строение органа слуха (рис. 2.1).
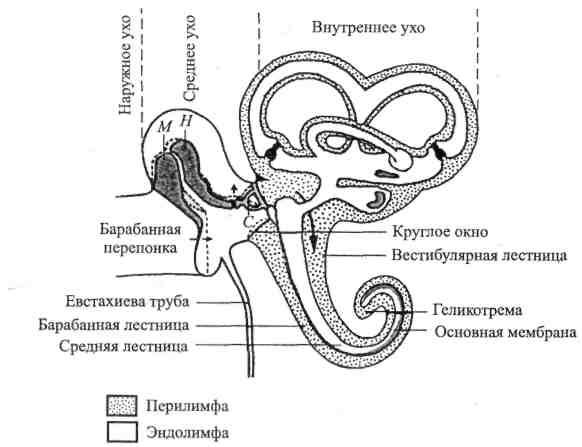
Рис. 2.1. Схема среднего и внутреннего уха (по Р. Шмидту и Г. Тевсу, 1985).
М – молоточек; Н – наковальня; С – стремечко. Пунктирной линией показаны крайние положения косточек под влиянием колебаний барабанной перепонки.
Звуковые волны направляются в слуховуюсистему через наружное ухо - наружный слуховой проход - к барабанной перепонке. Эта тонкая, с перламутровым блеском, мембрана замыкает слуховой проход, образуя преграду между ним и средним ухом, в котором также находится воздух.В полости среднего уха расположена цепочка подвижно сочлененных косточек: молоточек,наковальня истремечко, что делает более эффективной передачу звуковых колебаний из воздушного пространства наружного слухового прохода в жидкую среду внутреннего уха. Энергия звука передается во внутреннее ухо от барабанной перепонки через молоточек, наковальню и стремечко, колеблющиеся синхронно с ней. «Ручка» молоточка прочно связана с барабанной перепонкой, а основание стремечка (которое действительно выглядит как стремя) помещено в отверстие каменистой части височной кости -овальное окно (здесь стремечко граничит с внутренним ухом). Размеры площади овального окна значительно (в 22 раза) меньше воспринимающей поверхности барабанной перепонки, что способствует увеличению силы колебаний барабанной перепонки. Такое различие в размерах эффективной поверхности мембраны овального окна и барабанной перепонки, а также специальная система рычагов, образованная слуховыми косточками, создают условия для роста давления, прилагаемого к овальному окну, которое примерно в 20-22 раза больше давления, действующего на барабанную перепонку. В результате этого даже слабые звуковые волны, действующие на барабанную перепонку, способны преодолеть сопротивление мембраны овального окна преддверия и привести к колебаниям жидкости в улитке.
Полость среднего уха соединяется с глоткой посредством евстахиевой трубы, которая при глотании открывается, вентилируя среднее ухо и уравнивая в нем давление с атмосферным. Это особенно важно при резком перепаде давления - при взлете или посадке самолета, в метро во время начала движения поезда, в скоростном лифте или при близкой артиллерийской стрельбе, когда возникает состояние «закладывания ушей». В таком случае необходимостараться путем повышения давления в полости рта(открыть рот) или глотания уравнять его с возрастающим наружным давлением. В противном случае если это не удается, возникает опасность разрыва барабанной перепонки.
Внутреннее ухо помещается в каменистой части височной кости вместе с органом равновесия и представленокостным лабиринтом, внутри которого залегаетперепончатый лабиринт, повторяющий форму костного. Между обоими лабиринтами имеется щелевидное пространство, заполненное перилимфой.Лабиринт состоит из преддверия, трех полукружных каналов и улитки.Слуховым органом является улитка, названная так из-за своей формы(у человека она образует 2,5 завитка, у кошки и собаки– 3, у лошади – 2). Улитка состоит из трех параллельных свернутых вместе каналов -барабанной лестницы, вестибулярной лестницы и средней лестницы. Первые два соединяются между собой у вершины улитки через маленькое отверстие - геликотрему. Они заполнены перилимфой, сходной по химическому составу с внеклеточной жидкостью и содержащей поэтому много ионов натрия.
Средняя лестница заполнена эндолимфой – жидкостью, богатой ионами калия и, стало быть, сходна с внутриклеточной средой. Пери- и эндолимфатические области улитки соединяются с соответствующими областями вестибулярного аппарата. Основание стремечка в овальном окне примыкает к перилимфатическому пространству вестибулярной лестницы; отверстие замыкаетсякольцевой связкой, так что перилимфа не может вытекать. В основании барабанной лестницы имеется еще одно отверстие, открывающееся в среднее ухо, - круглое окно. Оно также закрыто тонкой мембраной, предотвращающей вытекание перилимфы.
На рис. 2.2 показано поперечное сечение улитки. Вестибулярная лестница отделяется от средней лестницы рейснеровой мембраной, а основная (базилярная) мембрана разделяет среднюю и барабанную лестницы. Утолщение, проходящее вдоль основной мембраны, -кортиев орган, который содержит фонорецепторы, окруженныеподдерживающими (опорными) клетками. Выделяют два вида опорных и рецепторных клеток – наружные и внутренние.
Опорные клетки сходятся под острым углом друг к другу и образуют заполненный эндолимфой канал треугольной формы – внутренний туннель, в котором расположены нервные волокна, идущие от нейроновспирального ганглия. Они также представлены волосковыми клетками, которые имеют, однако, только стереоцилии; в рецепторах кортиева органа киноцилии редуцированы.
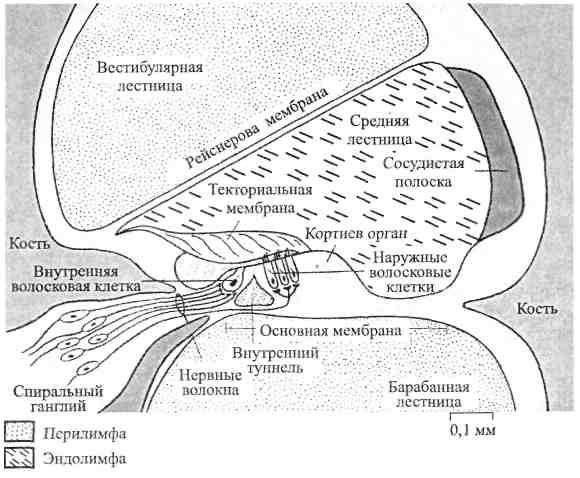
Рис. 2.2. Поперечный разрез одного из витков спирали улитки (по Р. Шмидту и Г. Тевсу, 1985).
Фонорецепторы лежат на опорных клетках. Как и в вестибулярном аппарате, рецепторные клетки здесь являются вторичными сенсорными клетками. На основании их отношения к кортиеву органу различают внутренние и наружные волосковые клетки; последние расположены в три ряда, тогда как внутренние образуют один. У человекаприблизительно 3500 внутренних и 12000 -20000 наружных волосковых клеток. На свободной поверхности фонорецепторов есть волоски, или стереоцилии (до 100 в клетке). Сегодня известно, что высокие звуки раздражают волосковые клетки, расположенные на нижних завитках улитки, а низкие звуки возбуждают волосковые клетки вершины улитки. Показано также, что внутренние волосковые клетки (в силу их более высокой чувствительности) лучше отражают интенсивность звука, чем наружные волосковые клетки.
Над кортиевым органом лежит текториальная (покровная) мембрана (имеет желеобразную консистенцию), один край которой прикрепляется к внутренней стенке улитки в области центральной кости, а другой свободно оканчивается в полости улиткового протока. Стереоцилии волосковых клеток связаны с нижней поверхностью текториальной мембраны.
Все фонорецепторы синаптически связаны с афферентными нервными волокнами,приходящим от биполярных клеток спирального ганглия, который расположенв центре улитки; центральные отростки этих клеток направляются в ЦНС. Около 90% нервных волокон спирального ганглия оканчиваются на внутренних волосковых клетках, каждая из которых образует контакты с множеством нервных волокон. Только оставшиеся 10% волокон иннервируют значительно более многочисленные наружные волосковые клетки. Чтобы охватить все наружные клетки, эти волокна должны сильно разветвляться, хотя иннервируемые одним волокном клетки могут быть расположеныблизко друг к другу. Всего слуховой нерв содержит примерно от 30 до 40 тысяч афферентных волокон. Кортиев орган получает и эфферентные волокна, тормозящие активность в афферентных волокнах.
На внешней стороне средней лестницы расположена сосудистая полоска с высокой метаболической активностью и хорошим кровоснабжением, чтои отражается в ее названии. Она играет важную роль вобеспечении улитки энергией и в регуляции состава эндолимфы. Различныеионные насосы, включая калиевый, поддерживают здесь постоянство ионной средыи положительный потенциал эндолимфы. Известно, что некоторые диуретические препараты являются ототоксичными и иногда вызывают глухоту. Это обусловлено тем, что указанные вещества воздействуют на почки, блокируя ионные насосы в эпителии канальцев, которые ответственны за реабсорбцию солей. Очевидно, некоторые из ионных насосов сосудистой полоски действуют по сходному принципу и, быть может, поэтому страдают при применении этих препаратов.
Механизмы слуховой рецепции.
Звуковая волна, воздействуя на систему слуховых косточек среднего уха, приводит в колебательное движение мембрану овального окна преддверия, которая, прогибаясь, вызывает волнообразные перемещения перилимфы верхнего (вестибулярной лестницы) и нижнего (барабанной лестницы) каналов, которые постепенно затухают по направлению к вершине улитки. Поскольку все жидкости несжимаемы, колебания эти были бы невозможны, если бы не мембрана круглого окна, которая выпячивается при надавливании основания стремечка на овальное окно и принимает исходное положение при прекращении давления. Колебания перилимфы передаются на вестибулярную мембрану, а также на полость среднего канала (среднюю лестницу), приводя в движение эндолимфу и базилярную мембрану (вестибулярная мембрана очень тонкая, поэтому жидкость в верхнем и среднем каналах колеблется так, как будто оба канала едины). При действии на ухо звуков низкой частоты (до 1000 Гц) происходит смещение базилярной мембраны на всем ее протяжении от основания до верхушки улитки. При увеличении частоты звукового сигнала происходит перемещение укороченного по длине колеблющегося столба жидкости ближе к овальному окну, к наиболее жесткому и упругому участку базилярной мембраны. Деформируясь, базилярная мембрана смещает стереоцилии волосковых клеток относительно текториальной мембраны (они наклоняются). Такое натяжение чисто механически открывает от 1 до 5 калиевых каналов в мембране стереоцилии, в результате чего возрастает поток ионов калия в стериоцилию (сила натяжения нити, необходимая для открывания одного канала, составляет всего около 2 • 10-13 ньютонов). Таким образом, в ответ на воздействие звука волосковая клетка деполяризуется, при этом максимальная величина рецепторного потенциала наблюдается уже спустя 100-150 мс от начала воздействия звука. Однако открывание всего нескольких ионных каналов в мембране одной стереоцилии недостаточно для возникновения рецепторного потенциала достаточной величины. Для усиления сенсорного сигнала на рецепторном уровне слуховой системы предусмотрен специальный механизм, который заключается в механическом взаимодействии всех стереоцилии каждой волосковой клетки. Дело в том, что все стереоцилии одного рецептора связаны между собой в пучок тонкими поперечными нитями. Поэтому, когда сгибается один или несколько более длинных волосков, они тянут за собой все остальные волоски. В результате этого открываются ионные каналы всех волосков, обеспечивая достаточную величину рецепторного потенциала. В свою очередь этот потенциал, т.е. деполяризация пресинаптического окончания волосковой клетки, приводит к выходу в синаптическую щель нейромедиатора. Воздействуя на постсинаптическую мембрану афферентного волокна, медиатор вызывает генерацию в нем возбуждающего постсинаптического потенциала и далее (при достижении критического уровня деполяризации) — генерацию распространяющихся в нервные центры импульсов.
►Проводящие пути слухового анализатора. Вся информация о звуке, воспринимаемая улиткой, передается в мозг волокнами преддверно-улиткового нерва (VIII пара черепных нервов). Существует пять основных уровней переключения восходящих слуховых волокон: кохлеарные ядра продолговатого мозга, верхнеоливарный комплекс ядер продолговатого мозга, нижние бугры крыши среднего мозга, медиальное коленчатое тело таламуса и слуховая зона коры больших полушарий (височные извилины). Кроме того, по ходу слухового пути расположено большое количество небольших ядер, в которых осуществляется частичное переключение восходящих слуховых волокон.
Выше уже отмечалось, что первыми нейронами слухового пути являются биполярные нейроны спирального ганглия, центральные отростки которых образуют слуховую, или кохлеарную, часть нерва — ветвь VIII пары черепно-мозговых нервов. По этому нерву информация от волосковых (главным образом, от внутренних) клеток поступает к нейронам продолговатого мозга, входящих в состав кохлеарного (улиткового) комплекса, т.е. к нейронам второго порядка. В этот комплекс, лежащий в области вестибулярного поля ромбовидной ямки, входят два ядра - дорсальное и вентральное (которое состоит из двух отделов — переднего и заднего). Аксон биполярного нейрона спирального ганглия, подходя к кохлеарным ядрам, раздваивается — одна его ветвь идет к дорсальному ядру, другая — к вентральному. Не исключено, что волокна, идущие от апикальной части улитки (т.е. несущие информацию о низких звуках) преимущественно достигают нейронов вентрального ядра, а волокна, идущие от основания улитки (возбуждаются высокими звуками) передают свою импульсацию, главным образом, нейронам дорсального ядра кохлеарного комплекса. Таким образом, для кохлеарных ядер характерно тонотопическое распределение информации.
Оба кохлеарных ядра дают восходящие тракты — дорсальный и вентральный. Аксоны нейронов дорсального кохлеарного ядра, не заходя к нейронам верхней оливы, сразу же направляются через мозговые полоски к латеральному лемниску, где часть из них переключается на нейроны лемниска (III нейроны), а часть проходит транзитом до нейронов нижних бугров четверохолмия или до нейронов медиального коленчатого тела.
Аксоны вентрального кохлеарного ядра сразу же направляются к варолиеву мосту через трапециевидное тело к верхней оливе, где находится верхнеоливарный комплекс (часть волокон идет к ипсилатеральному комплексу, часть — к контрлатеральному). В его состав входят два ядра: латеральное и медиальное. Медиальное ядро получает информацию одновременно как от ипсилатрального, т.е. со своей стороны, так и от контрлатерального, т.е. противоположной стороны, кохлеарного ядра, что обеспечивает формирование бинаурального слуха уже на уровне верхней оливы.
Аксоны верхнеоливарнных нейронов направляются к латеральному лемниску, где часть из них переключаются на нейроны этого лемниска (IV нейроны), а часть проходит транзитом к нейронам нижних бугров четверохолмия и к нейронам медального коленчатого тела, которое является последним переключательным звеном восходящего слухового пути.
Таким образом, от дорсальных и вентральных кохлеарных ядер информация в конечном итоге поступает к нижним буграм четверохолмия и медальному коленчатому телу. Благодаря этому звуковая информация используется для реализации ориентировочного рефлекса на внезапное звуковое раздражение (поворот головы в сторону источника звука), а также для регуляции тонуса скелетной мускулатуры и формирования взгляда. Одновременно, от нейронов медиального коленчатого тела информация достигает нейронов верхней части височной доли мозга (поля 41 и 42 по Бродману), т.е. высших акустических центров, где совершается корковый анализ звуковой информации.
Следует подчеркнуть, что для верхнеоливарного комплекса, нижних бугров четверохолмия, медиального коленчатого тела, а также для первичных проекционных зон слуховой коры, т.е. для всех важнейших слуховых центров характерна тонотопическая организация структур. Это отражает существование принципа пространственного анализа звуков, позволяющего осуществлять тонкое частотное различение на всех этажах слуховой системы.
Чрезвычайно важным свойством слуховой системы является билатеральная иннервация структур на каждом уровне. Впервые она появляется на уровне верхней оливы и дублируется на каждом последующем уровне. Это позволяет реализовать способность человека и животных оценивать место расположения источника звука.
Наряду с восходящими путями в слуховой системе имеются и нисходящие пути, обеспечивающие контроль высших акустических центров над получением и обработкой информации в периферическом и проводниковом отделах слухового анализатора. Нисходящие пути слухового анализатора начинаются от клеток слуховой коры, переключаются последовательно в медиальных коленчатых телах, нижних буграх четверохолмия, верхнеоливарном комплексе, от которого идет оливокохлеарный пучок Расмуссена, достигающий волосковых клеток улитки. Кроме того, имеются эфферентные волокна, идущие от первичной слуховой зоны, т.е. от височной области, к структурам экстрапирамидной двигательной системы (базальным ганглиям, ограде, верхним буграм четверохолмия, красному ядру, черной субстанции, некоторым ядрам таламуса, ядрам основания моста, ретикулярной формации ствола мозга) и пирамидной системы. Эти данные указывают на участие слуховой сенсорной системы в регуляции двигательной активности человека.
►Переработка информации в коре больших полушарий. В повседневной жизни мы почти никогда не сталкиваемся с чистыми тонами. Звуки, которые мы слышим, построены из разных частотных компонентов, которые постоянно и независимо меняются (человек воспринимает звуковые колебания с частотой 16-20000 Гц). Амплитуда таких звуков также может меняться, как и их частота, варьирует и их длительность, они также могут начинаться и заканчиваться резко и постепенно, их источник может располагаться ближе, дальше или перемещаться и т.д. Конечно, в полном объеме люди способны оценивать все эти свойства звука, только если их слух достаточно тренирован. Так вот, нейронные процессы, лежащие в основе такой оценки, выявляются в основном в слуховой области коры (средняя часть верхней височной извилины). Так, например, одни нейроны первичной слуховой коры отвечают только на начало звукового стимула, а другие – только на его окончание. Некоторые другие возбуждаются только при звуках определенной длительности либо при повторяющихся звуках. Есть нейроны, возбуждение которых возникает только в случае, если стимул определенным образом модулирован по частоте или амплитуде. Многие нейроны активируются при действии звуков в широкой полосе частот, т.е. шумов, а другие, так называемые простые нейроны, — вычленяют информацию о чистых звуках. Есть нейроны, которые возбуждаются только на определенную последовательность звуков или на определенную амплитудную их модуляцию. Есть нейроны, которые позволяют определить направление звука. Повреждение височных долей мозга, т.е. именно тех, где располагается слуховая кора, затрудняет восприятие речи, пространственную локализацию источника звука и идентификацию временных характеристик звука. Подобные повреждения, однако, не влияют на способность различать частоты и интенсивность звука (см. выше).
Большинство корковых нейронов активируется контралатеральным ухом, некоторые отвечают на ипсилатеральную стимуляцию, а остальные – только при одновременной стимуляции того и другого уха. Значительный процент нейронов первичной слуховой коры не возбуждаются ни при каких экспериментальных воздействиях. Предполагается, что эти нейроны являются высокоспецифичными и отвечают только на стимулы, которые сложно воспроизвести в лабораторных условиях. В целом ответы клеток первичной слуховой коры сходны с ответами групп или ансамблей нейронов зрительной коры.
В целом, в первичных и вторичных проекционных зонах слуховой коры происходит сложнейший анализ звукового сигнала. Однако важной является и функция ассоциативных зон коры больших полушарий. Например, представление о мелодии возникает именно благодаря деятельности этих зон коры, в том числе на основе информации, хранящейся в памяти. Именно с участием ассоциативных зон коры (с помощью специализированных нейронов типа «бабушкиных» нейронов) человек способен максимально извлечь информацию, поступающую от различных рецепторов, в том числе и от фонорецепторов.