26.Промотор прокариот и механизм его распознавания рнк-полимеразой. Альтернативные s-факторы(этого калла нет, но есть не s факторы а сигма, что и описаны ниже). Стадии транскрипционного цикла.


Альтернативные сигма-факторы играют важную роль в регуляции генной экспрессии у прокариот. Они позволяют РНК-полимеразе распознавать различные промоторы и тем самым инициировать транскрипцию разных генов в ответ на изменения в окружающей среде.
Сигма-факторы являются подразделением белковой подединицы РНК-полимеразы, которая связывается с промотором ДНК и инициирует процесс транскрипции.
Прокариотическая РНК-полимераза распознает определенные последовательности ДНК, называемые промоторами, которые отмечают начальные точки транскрипции. Промоторы содержат консенсусные последовательности, которые распознаются РНК-полимеразой и связанными с ней факторами.
У прокариот промотор включает ряд консервативных мотивов, важных для узнавания его РНК-полимеразой, в частности так называемые последовательности -10 и -35. Промотор асимметричен, что позволяет РНК-полимеразе начать транскрипцию в правильном направлении и указывает то, какая из двух цепей ДНК будет служить матрицей для синтеза РНК.
Во время транскрипции РНК-полимераза связывается с определенной областью ДНК, называемой промотором. Это взаимодействие запускает раскручивание двойной спирали ДНК, позволяя ферменту получить доступ к цепи матрицы. Затем РНК-полимераза начинает синтезировать РНК1. РНК-полимераза функционирует в сотрудничестве с различными белками для осуществления транскрипции. Эти белки способствуют специфическому связыванию РНК-полимеразы с ДНК, облегчают раскручивание нитей ДНК, модулируют ферментативную активность РНК-полимеразы и регулируют скорость и точность транскрипции
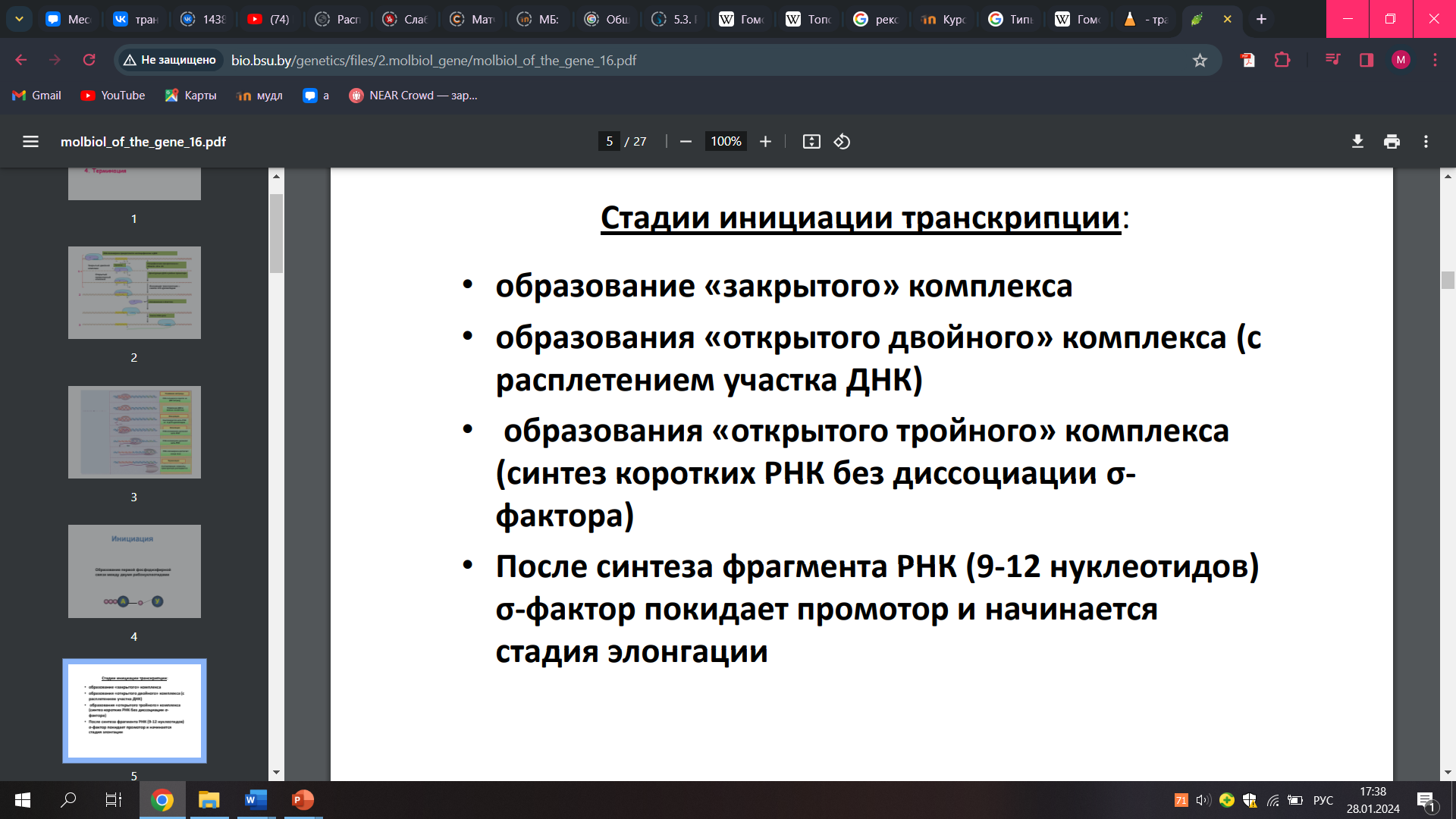
В процессе следующей стадии – элонгации – происходит удлинение цепи РНК. При этом новосинтезированная цепь РНК образует короткие отрезки гибридной двойной спирали ДНК-РНК (рис. 4.2), которые необходимы для правильного копирования матричной цепи ДНК.
Как только РНК-полимераза приблизится к терминатору, начинается последняя стадия – терминация, в результате которой происходит распад тройного комплекса и освобождение вновь синтезированной молекулы РНК – транскрипта.
27.Промоторы эукариот: размеры, положение, структура и механизм
распознавания различными РНК-полимеразами.
Промотор представляет собой последовательность нуклеотидов, обеспечивающую базальный (но не максимальный) уровень транскрипции соответствующего транскриптона. Он является той минимальной последовательностью, которая специфически распознается холоферментом РНК-полимеразы среди случайных последовательностей нуклеотидов.
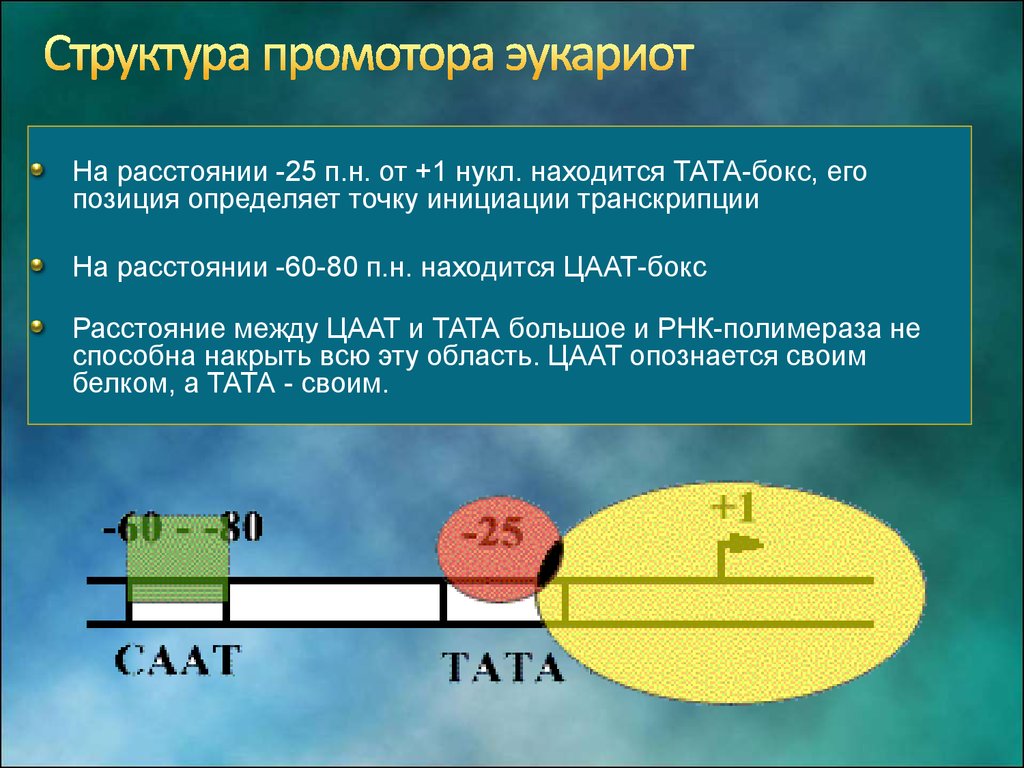
Промотор эукариотических генов (размер 40-60 п.н.), узнающийся РНК-полимеразой промотор содержит два базовых регуляторных элемента: ТАТА-последовательность (положение -25) и специфическую нуклеотидную последовательность, обогащенную пиримидинами в положении -75.
Основной элемент промотора - место связывания РНК-полимеразы, которое она занимает перед началом синтеза РНК. В состав промоторов могут входить также участки связывания белков-регуляторов.
Механизм У эукариот регуляторные элементы собраны в регуляторные регионы. Основной регуляторный элемент эукариот – это коровый (базальный) промотор. Он обеспечивает сборку базального транскрипционного комплекса (из основных факторов транскрипции и РНК-полимеразы) и инициацию транскрипции на базальном (исходном, базовом) уровне. Часто этот уровень так низок, что приводит к синтезу лишь единичных молекул РНК и, в дальнейшем, белков.
Промоторные элементы, контролирующие точку инициации и интенсивность транскрипции.
Промоторы, узнаваемые РНК-полимеразой II, содержат три различных семейства регуляторных последовательностей ДНК. Последовательности первого семейства включают так называемые базальные элементы промотора, расположенные вблизи точки инициации транскрипции. В настоящее время известны два класса базальных элементов: TATA-последовательность, расположенная за 25–30 нуклеотидов до точки инициации (каноническая последовательность – TATAa/tAa/t), и так называемый инициатор (Inr), последовательность которого обогащена пиримидинами. Элементы TATA-последовательности и инициатор необходимы для сборки ДНК-белкового инициационного комплекса и распознаются основными факторами транскрипции. Промоторы РНК-полимеразы II содержат один или оба регуляторных элемента или же не имеют их вообще. При этом оба элемента могут функционировать независимо друг от друга или же в их действии наблюдается синергизм.
|
К двум другим классам цис- регуляторных промоторных элементов у эукариот относятся последовательности, расположенные вблизи промотора (от 50 до нескольких сотен пар оснований перед точкой инициации), а также дистальные элементы (энхансеры и сайленсеры), расстояние которых от промотора может превышать 60 т.п.о. Оба класса таких последовательностей содержат сайты связывания регуляторных белков, модулирующих транскрипцию.