

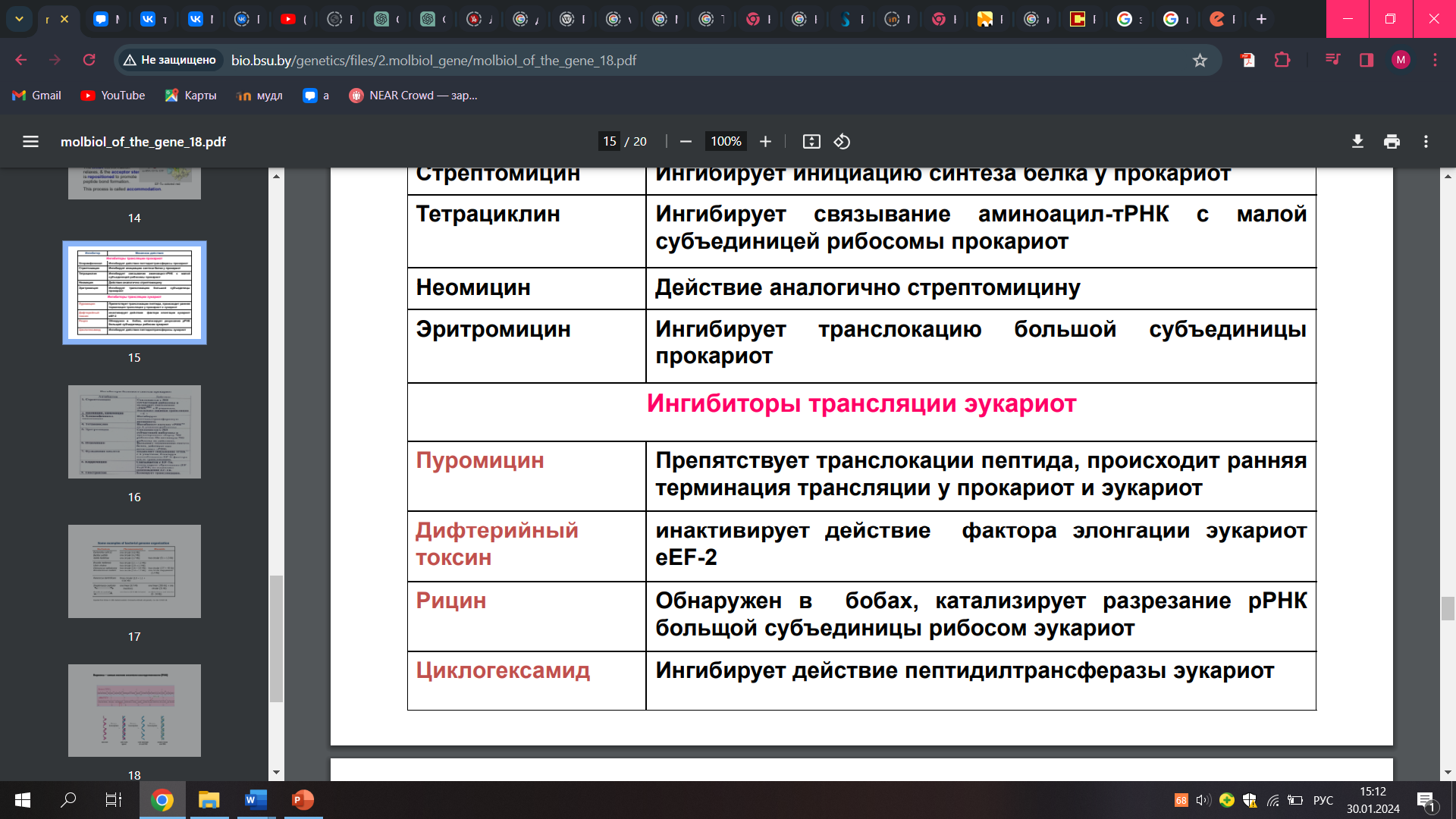
44. Ингибиторы синтеза белка

45. Молекулярные шапероны семейства Hsp60. Рабочий цикл шаперонина GroEls
Белки Hsp70 и Hsp60 называют белками теплового шока (heat shock proteins), поскольку они активно синтезируются клеткой при нагревании. Гомологом Hsp60 у E. coli является GroELS.
GroE (GroEL/GroES) — шаперонин бактерий. Шаперонины— это функциональная группа белков, которая входит в более широкий класс белков — шапероны. Шапероны ассистируют другим белкам при их синтезе, транспорте, деградации. Самая известная функция шаперонов — помогать только что синтезированным или поврежденным белкам обрести правильную пространственную структуру.
GroE представляет собой мультисубъединичный комплекс (состоящий из множества мономеров — субъединиц), похожий на горшочек с крышкой. «Горшочек» — шаперонин GroEL — образован двумя кольцами, лежащими друг на друге. Каждое такое колечко — гептамер, то есть состоит из семи одинаковых белковых субъединиц. Субъединицы в обоих кольцах одинаковые, но находятся в разной конфигурации. «Крышка» (красная на верхнем рисунке) — GroES — это тоже гептамерный кольцевой комплекс, но мономеры в нем другие.
Внутри комплекса Есть полость, в которую попадают Белки, они узнаются шаперонином по торчащим гидрофобным неполярным участком, в норме они не выставлены в цитоплазму, а находятся внутри белковой структуры.
Торчащие гидрофобные участки, это как раз то, что приводит к агрегации не правильно сложенных белков, когда субъединицы кольца GROEL с помощью специально расположенных гидрофобных остатков ловят неправильно сложенный белок, каждая из 7ми субъединиц связывает молекула АТФ с цитоплазмы.
Конформация комплекса меняется, его внутреннее пространство расширяется, что способствует разворачиванию неправильно структуры пойманного белка. Кольцо получает способность соединиться с крышкой GROES, которая утрамбовывает белок. Присоединение GROES снова меняет конформацию субъединиц кольца GROEL. Окружение белка из гидрофобного становился гидрофильным, что может навести его на принятие правильной структуру. При такой конформации комплекса, начинает происходить гидролиз связанных их молекул атф, что необходимо для открытия крышки и высвобождение белка
46. Классы генов теплового шока у b. Subtilis. Рабочий цикл шаперонного комплекса DnaKj-GrpE
У B. subtilis гены теплового шока могут быть разделены на три класса:

DnaKJ-GrpE
У E. coli гомологом Hsp70 является DnaK, который образует комплекс с кошапероном DnaJ и белком GrpE
В активный комплекс с Hsp70 входят белки-ко-шапероны Hsp40 и Hsp10. Ко-шаперон Hsp40 (мол. масса 40 кДа, в E. coli – DnaJ) содержит консервативный J домен (70 аминокислотных остатков), который определяет контакт с Hsp70. Кроме того, DnaJ содержит G/F-богатый фрагмент, цинксвязывающий домен и С-терминальный домен, участвующий в контакте с субстратом. Третий белок - Hsp10, необходимый для проявления шаперонной активности комплекса Hsp70-Hsp40 (DnaK-DnaJ), в E. coli, – GrpE. GrpE способствует быстрому обмену АТФ – АДФ, ускоряя освобождение комплекса от АДФ. Кратко данную систему шаперонов в E. coli обычно обозначают как DnaKJE.

![]()
Ко-шаперон GrpE необходим для диссоциации АДФ и для усиления связи с АТФ, в результате происходит высвобождение ренатурированного субстрата из комплекса и завершается цикл действия шаперонной системы