

28.Регуляция процесса транскрипции прокариот. Лактозный и триптофановый опероны. Про опероны (изучите как они работают в различных ситуациях, здесь такого нет!!!!)
Лактозный оперон – данный оперон кодирует белки катаболизма лактозы, менее предпочтительного по сравнению с глюкозой источника энергии для данной бактерии. В связи с этим транскрипция lac-оперона происходит только в том случае, если в среде отсутствует глюкоза и присутствует лактоза. В состав lac-оперона входят: три структурных гена, кодирующих белки, участвующих в метаболизме лактозы: βгалактозидазу (гидролизует лактозу), пермеазу (ответственна за транспорт галактозы в клетку), белок А (β-галактозидтрансацетилаза – функция до конца не выяснена); промотор (40 п.н.); оператор (27 п.н.); область связывания комплекса белка САР (белок активатор катаболитических оперонов) с цАМФ – САР•цАМФ (38 п.н.).
+Триптофановый оперон содержит 5 генов, ответственных за синтез триптофана, промотор, оператор.
Триптофановый оперон должен транскрибироваться при недостаточном содержании триптофана в клетке. При высоком же содержании этой аминокислоты его экспрессия должна быть слабо выраженной или полностью отсутствовать. В клетках E.coli присутствует репрессор, образующийся в результате экспрессии соответствующего гена. При высоком содержании в клетке триптофана этот репрессор образует комплекс с данной аминокислотой. Образовавшийся комплекс обладает сродством к оператору и, взаимодействуя с ним, не позволяет РНК-полимеразе начать синтез РНК. Белки, ответственные за синтез триптофана, не образуются. Соответственно не происходит и синтез аминокислоты.
Как только содержание триптофана в клетке снижается, комплекс триптофан-репрессор распадается. В результате репрессор теряет сродство к оперону. РНК-полимераза приобретает доступ к промотору и начинает транскрипцию структурных генов. Синтезируется иРНК, которая транслируется с образованием белков, ответственных за синтез триптофана. Концентрация этой аминокислоты в клетке постепенно возрастает.
29. Нематричный синтез рнк.
 Существуют
ферменты способные синтезировать РНК
в отсутствие матрицы. К ним относится
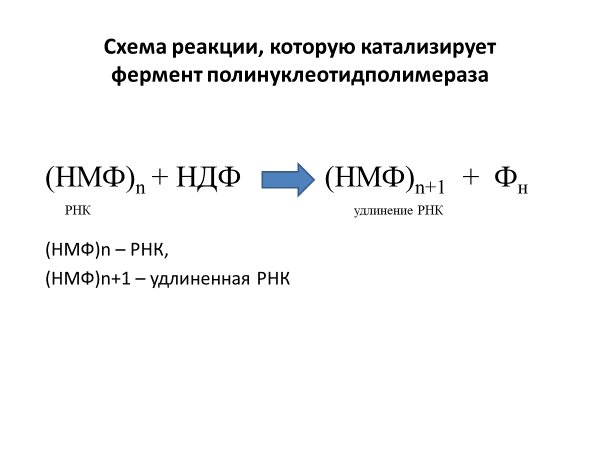
полинуклеотидполимераза и
полиаденилатполимераза. Полинуклеотидполимераза
– встречается у бактерий и использует
в качестве субстратов рибонуклеозид-5’-дифосфаты
(НДФ) – АДФ, ЦДФ, ГДФ и УДФ. Этот фермент
катализирует реакцию, которую в общем
виде можно представить так:
Существуют
ферменты способные синтезировать РНК
в отсутствие матрицы. К ним относится
полинуклеотидполимераза и
полиаденилатполимераза. Полинуклеотидполимераза
– встречается у бактерий и использует
в качестве субстратов рибонуклеозид-5’-дифосфаты
(НДФ) – АДФ, ЦДФ, ГДФ и УДФ. Этот фермент
катализирует реакцию, которую в общем
виде можно представить так:
Реакция обратима.. Полиаденилатполимераза обеспечивает синтез полиА-последовательности на 3’-конце иРНК. В качестве субстрата использует АТФ и РНК..
30. Информационная рнк, ее структура и функциональные участки, различия у про-и эукариот. Модификация 5'- и 3'-концов транскриптов и ее значение.
иРНК (или матричные РНК) – это одноцепочечные молекулы. Они содержат информацию о первичной структуре белка и служат матрицами для биосинтеза полипептидных цепей.
Молекула иРНК может кодировать одну или несколько полипептидных цепей. иРНК, кодирующая информацию об одной полипептидной цепи, называется моноцистронной, о двух или более – полицистронной.
Информационная РНК прокариот
иРНК
прокариот – это короткоживущие молекулы,
их время полужизни составляет всего
около 2 мин. иРНК прокариот в большинстве
случаев являются полицистронными, и
только некоторые из них – моноцистронными.
Полицистронные иРНК содержат
нетранслируемые межцистронные области,
которые разделяют участки, кодирующие
полипептидные цепи (цистроны). Со стороны
5’ располагается лидерная не кодирующая
последовательность, содержащая в своем
составе Шайна-Дальгарно, определяет
правильную ориентацию на рибосомах, со
стороны 3’ кольцевая последовательность
- трейлер. 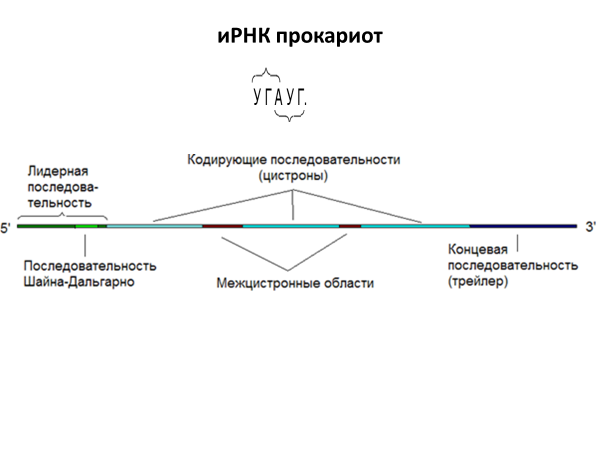
иРНК эукариот
иРНК эукариот являются моноцистронными . На их 5’-конце имеется кэп. Кэп представляет собой 7-метилгуанозин, связанный трифосфатной связью 5’-5’со следующим нуклеотидом. Присоединение кэпа происходит посттранскрипционно. Кэп играет важную роль в инициации синтеза белка.
 На
3’-конце большинства иРНК эукариот
имеется полиАпоследовательность. ПолиА-
последовательность не закодирована в
генах, поэтому полиаденилирование
осуществляется после транскрипции (в
ядре) ферментом полиаденилатполимеразой.
ПолиА-последовательность имеет размер
около 50 – 400 нуклеотидов. В цитоплазме
она постепенно укорачивается и определяет
время жизни иРНК. Таким образом, эта
последовательность защищает иРНК от
деградации последовательности. Кроме
того, она способствует транспорту иРНК
из ядра в цитоплазму. На 5’-и 3’- флангах
иРНК располагаются нетранслируемые
области. Период полужизни иРНК эукариот
значительно больше, чем у прокариот, и
составляет от нескольких часов до
нескольких суток.
На
3’-конце большинства иРНК эукариот
имеется полиАпоследовательность. ПолиА-
последовательность не закодирована в
генах, поэтому полиаденилирование
осуществляется после транскрипции (в
ядре) ферментом полиаденилатполимеразой.
ПолиА-последовательность имеет размер
около 50 – 400 нуклеотидов. В цитоплазме
она постепенно укорачивается и определяет
время жизни иРНК. Таким образом, эта
последовательность защищает иРНК от
деградации последовательности. Кроме
того, она способствует транспорту иРНК
из ядра в цитоплазму. На 5’-и 3’- флангах
иРНК располагаются нетранслируемые
области. Период полужизни иРНК эукариот
значительно больше, чем у прокариот, и
составляет от нескольких часов до
нескольких суток.