

31. Интроны. Особенности структуры и механизмы сплайсинга. Аутосплайсинг.
Многие гены эукариот состоят из экзонов – кодирующих последовательностей и интронов – некодирующих последовательностей. При транскрипции таких генов считывается РНК, содержащая в своем составе как экзоны, так и интроны. Образовавшийся первичный транскрипт, подвергается процессингу, в результате которого интроны вырезаются, а экзоны, сшиваясь, образуют зрелую РНК. Данный процесс получил название сплайсинг.

Сплайсингу подвергаются предшественники различных эукариотических РНК: иРНК, тРНК, рРНК.
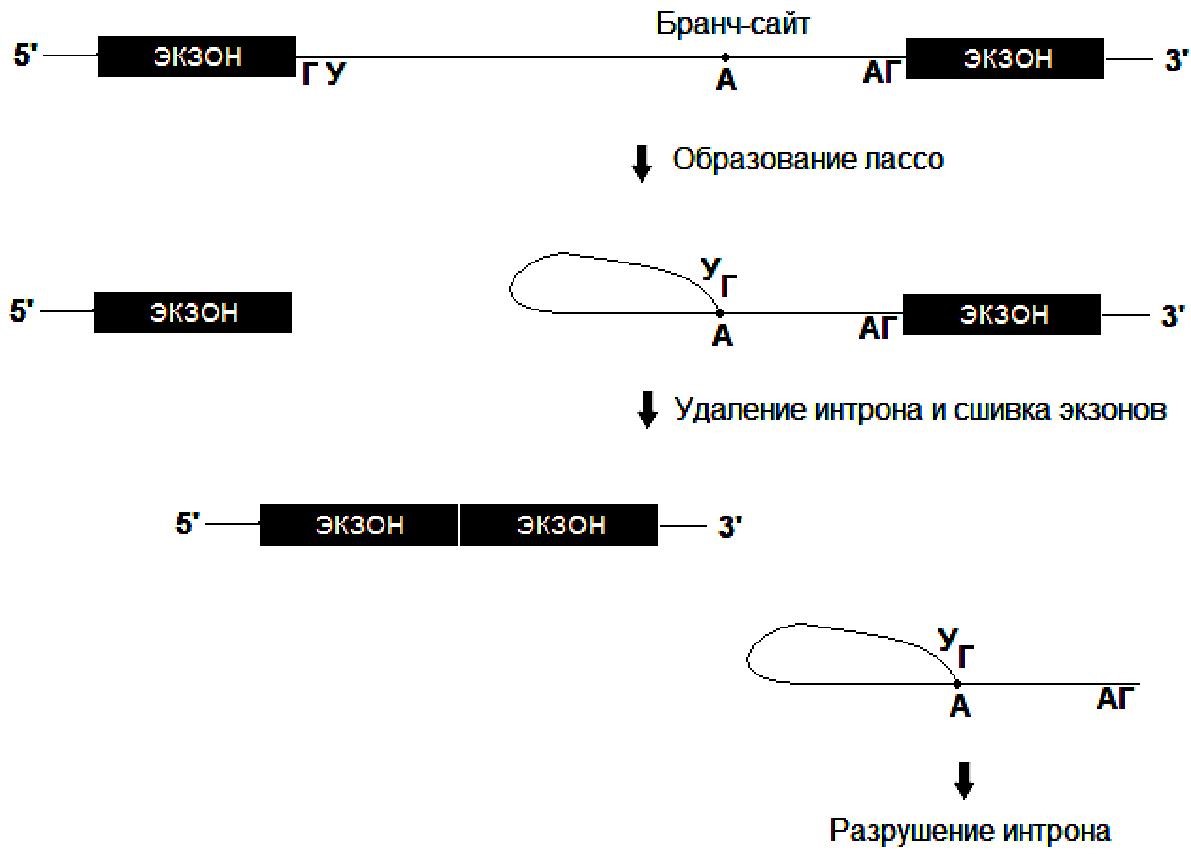
У эукариот в предшественниках иРНК (пре-РНК) интроны вырезаются в сплайсосомах – комплексах, состоящих из мяРНК и белков. Интроны на границах с экзонами имеют канонические последовательности на 5’-конце – ГУ, на 3’-конце – АГ. Они также содержат последовательности, необходимые для удаления: акцепторный сайт, донорный сайт и бранч-сайт

Процесс удаления интронов включает:
a) разрыв молекулы РНК на границе интрон-экзон со стороны 5’-конца интрона;
b) образование сложнофирных связей между фосфатной группой первого нуклеотида интрона (Г) и гидроксильной группой рибозы (у 2’ атома углерода) аденозина, входящего в состав бранч-сайта. Сформировавшаяся структура напоминает лассо;
c) удаление и последующее разрушение интрона;
d) воссоединение экзонов посредством образования фосфодиэфирных связей.
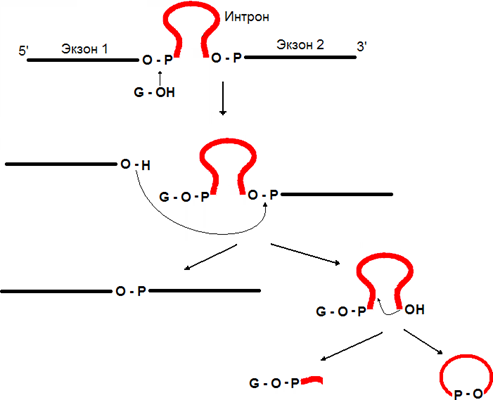
Аутосплайсинг (самосплайсинг) – сплайсинг первичного транскрипта РНК, происходящий без участия каких-либо ферментов, т.е. РНК сама является катализатором этого процесса. Для протекания сплайсинга необходимо присутствие нуклеотида содержащего гуанин (ГТФ, или ГДФ, или ГМФ, или гуанозин). На 3’-гидроксильную группу этого гуанинового нуклеотида переносится фосфатная группа 5’-конца интрона. Затем образовавшаяся на конце первого экзона 3’-гидроксильная группа используется для присоединения к 5’-концу второго экзона посредством фосфодиэфрной связи. Вырезание интрона сопровождается его циклизацией и удалением из его состава небольшого фрагмента, содержащего то гуаниновый нуклеотид, который использовался для инициации сплайсинга.
32. Сплайсинг пре-тРнк.
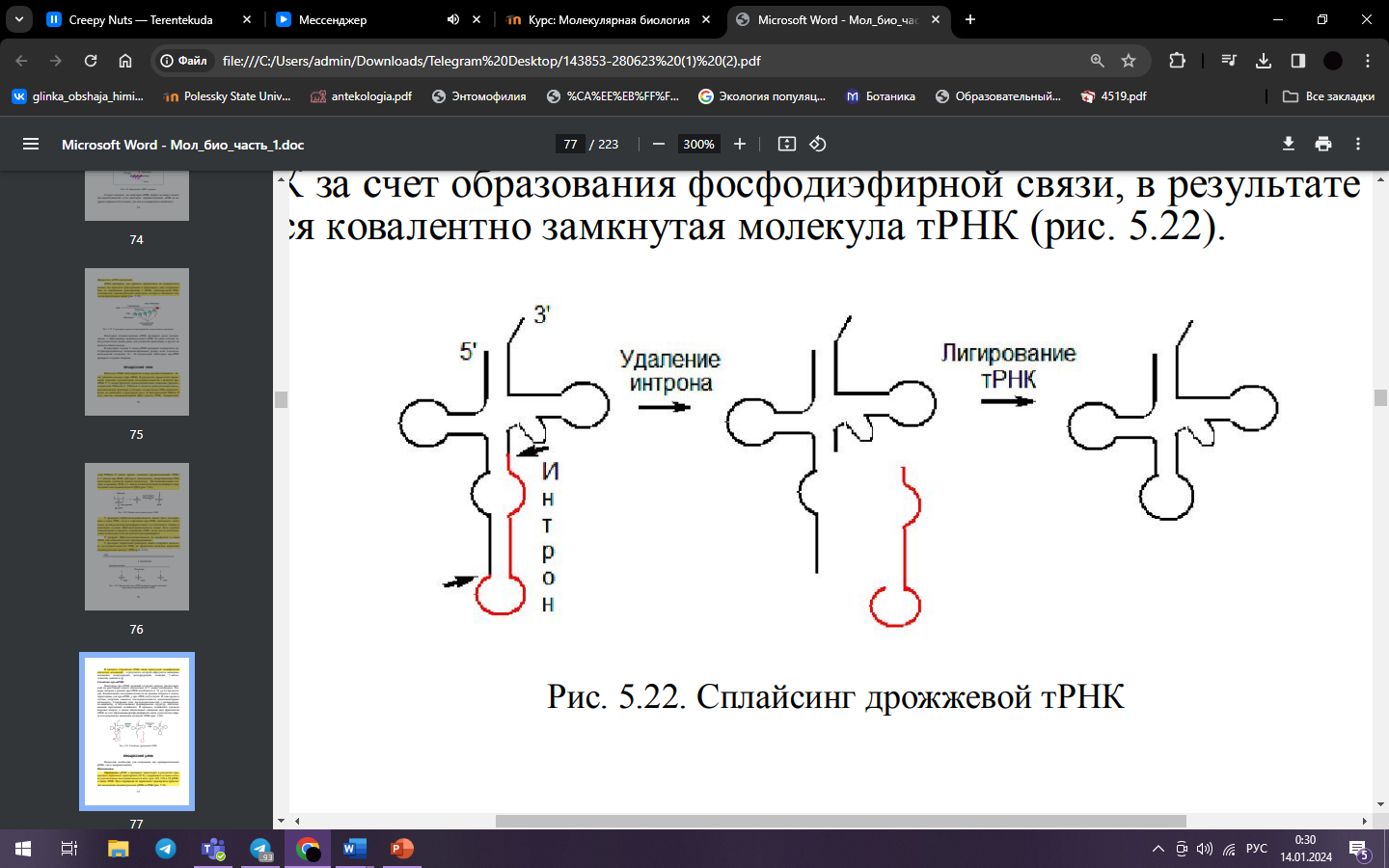 Некоторые
пре-тРНК дрожжей содержат интрон,
расположенный на расстоянии одного
нуклеотида от 3’-конца антикодона.
Размеры интрона у разных пре-тРНК
колеблются от 14 до 64 нуклеотидов.
Канонические последовательности на
границе интрона и экзона, характерные
для пре-иРНК, у пре-тРНК отсутствуют. В
тоже время в составе интронов имеются
последовательности комплементарные
антикодону. Спаривание этих
последовательностей с антикодоном,
по-видимому, и обуславливает формирование
структур, обеспечивающих протекания
сплайсинга. В процессе сплайсинга
нуклеаза вырезает интрон, а лигаза
обеспечивает сшивание двух фрагментов
тРНК за счет образования фосфодиэфирной
связи, в результате образуется ковалентно
замкнутая молекула тРНК.
Некоторые
пре-тРНК дрожжей содержат интрон,
расположенный на расстоянии одного
нуклеотида от 3’-конца антикодона.
Размеры интрона у разных пре-тРНК
колеблются от 14 до 64 нуклеотидов.
Канонические последовательности на
границе интрона и экзона, характерные
для пре-иРНК, у пре-тРНК отсутствуют. В
тоже время в составе интронов имеются
последовательности комплементарные
антикодону. Спаривание этих
последовательностей с антикодоном,
по-видимому, и обуславливает формирование
структур, обеспечивающих протекания
сплайсинга. В процессе сплайсинга
нуклеаза вырезает интрон, а лигаза
обеспечивает сшивание двух фрагментов
тРНК за счет образования фосфодиэфирной
связи, в результате образуется ковалентно
замкнутая молекула тРНК.
33. +-Сплайсинг пре-мРНК в ядре(в инете пишут что идет по альетнативному сплайсингу). Принципы определения границ интронов у разных организмов(вспомните 32 вопрос и 31). Сплайсосома (размеры и состав). мяРНК и мяРНП-частицы. Роль комплементарных взаимодействий в протекании процесса сплайсинга(это вообще хз, но есть что-то)
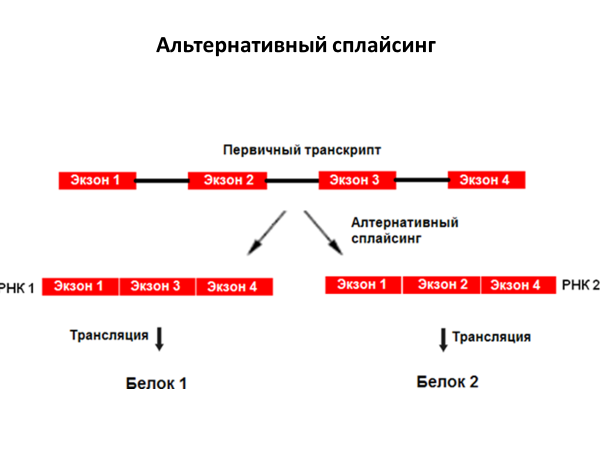 Альтернативный
сплайсинг обеспечивает образования
разнообразных белков с одного и того
же первичного транскрипта.
Альтернативный
сплайсинг обеспечивает образования
разнообразных белков с одного и того
же первичного транскрипта.
Точное определение границ интронов зависит от нескольких консервативных последовательностей в их составе. Для интронов пре-мРНК это донорный и акцепторный сайты сплайсинга, точка ветвления и пиримидиновый тракт.
Сплайсосомы – комплексы, состоящие из мяРНК и белков. Сплайсосома состоит из пяти рибонуклеопротеиновых U-частиц, молекулы РНК которых за счет комплементарных взаимодействий обеспечивают формирование каталитического центра сплайсосомы и распознавание концов интронов.
Сплайсинг РНК катализируется комплексами белков с РНК, известными как «малые ядерные рибонуклеопротеидные частицы» (мяРНП, англ. small nuclear ribonucleic particles, snRNP).
Малые ядерные РНК (мяРНК) – короткие молекулы, принимающие участие в созревании РНК. Их размер составляет от 65 до 1000 и более нуклеотидов.
Комплементарные взаимодействия, такие как спаривание вторичной структуры РНК, внутренние итерационные петли, базовые взаимодействия и специфичность взаимодействия белков, играют решающую роль в определении точек резания (splice sites) и мест соединения экзонов в процессе сплайсинга.
Они служат для распознавания и определения правильных спайс-сайтов, поскольку в геноме может быть много потенциальных спайс-сайтов, и необходимо точно определить, какие участки должны быть удалены и какие должны быть соединены для правильного образования зрелой мРНК.