

1 курс / Гистология / Общий курс гистологии
.pdfв экваториальной плоскости веретена (экваториальная пластинка). Анафаза длится 2–3 мин, дочерние хромосомы при участии полярных микротрубочек веретена смещаются к противоположным полюсам. Телофаза длится 30–40 мин, формируются оболочка дочерних ядер, эухроматиновые и гетерохроматиновые области, ядрышки, происходит цитотомия.
Мейоз: профаза I → метафаза I → анафаза I → телофаза I → прометафаза II → метафаза II → анафаза II → телофаза II (без синтеза ДНК в промежутке между I и II делениями и образованием 4-х гаплоидных клеток с новой комбинацией генов).
В мейозе 90% времени занимает профаза 1-го деления, поэтому в ней, в свою очередь, выделяют 5 стадий: лептонему, зиготему, пахитему, диплонему и диакинез. Лептонема – стадия тонких светооптически различимых хромосом. Зиготема – конъюгация гомологичных хромосом в пары. Пахитема – стадия толстых хромосом, перекреста и обмена участками. Диплонема – частичное расхождение дочерних хромосом, образование тетрад. Диакинез – отделение от оболочки ядра генетически измененных хромосом, состоящих из 2-х хроматид. В анафазу I расходятся к полюсам гомологичные хромосомы, анафазу II
– хроматиды: 2n2с → 2n4с (тетраплоидная клетка в S-период перед 1- м делением) → 1n2с (2 диплоидные клетки после 1-го деления) → 1n1с (4 гаплоидные клетки после 2-го деления).
Гибель клеток осуществляется генетически запрограммированным способом (апоптоз) и происходит при значительном поврежде-
нии клеток (некроз). |
|
|
|
|
|
|
|
|
||||
При апоптозе, благо- |
|
|
|
|
|
|
|
|
||||
даря |
экспрессии |
особых |
|
|
|
|
|
|
|
|
||
генов, инициируемых раз- |
|
|
|
|
|
|
|
|
||||
личными факторами, в том |
|
|
|
|
|
|
|
|
||||
числе и неблагоприятными |
|
|
|
|
|
|
|
|
||||
|
|
|
|
1 |
|
|
|
|||||
для клетки, активизируется |
|
|
|
|
|
|
|
|||||
|
2 |
|
|
|
|
2 |
|
|||||
ферментативный |
каскад. |
|
|
|
|
|||||||
При |
участии |
ферментов |
|
|
|
|
|
|
|
|
||
фрагментируются |
хрома- |
|
|
|
|
|
|
|
|
|||
тин и цитоскелет, поверх- |
|
|
|
|
|
|
|
|
||||
ность ядра и клетки стано- |
|
|
|
|
|
|
|
|
||||
вятся складчатыми, хрома- |
|
|
|
|
|
|
|
|
||||
Рис. 4. Апоптоз секреторного нейрона гипота- |
||||||||||||
тин конденсируется в про- |
||||||||||||
ламуса крысы: |
1 – апоптотическое ядро, 2 – |
|||||||||||
туберанцах |
апоптотиче- |
|||||||||||
апоптотические почки. Электронная микрофото- |
||||||||||||
ского ядра в виде крупных |
||||||||||||
графия, ув. ×10000 (препарат А.В. Герасимова) |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
глыбок. Затем апоптотические почки преобразуются в ограниченные мембраной апоптотические тельца (рис. 4).
Некроз: нарушение трофики → дефицит АТФ → аутолиз (нейтральный красный диффузно окрашивает цитоплазму, в которой из-за снижения рН и активации лизосомных ферментов исчезают барьеры), ядро сморщивается (кариопикноз), фрагментируется (кариорексис), рассасывается (кариолизис) – «ядерная тень» → «клеточная тень» (детрит).
Плазмолемма – наружная клеточная мембрана толщиной 10 нм. В электронном микроскопе в ней различимы 5 структурных компонентов: наружная и внутренняя плотные, промежуточная прозрачная пластинки, внутримембранные частицы и гликокаликс. Наружная плотная пластинка образована гидрофильными «головками» фосфолипидов наружного слоя. Внутренняя плотная пластинка образована гидрофильными «головками» фосфолипидов внутреннего слоя. Промежуточная прозрачная пластинка образована гидрофобными «хвостами» фосфолипидов наружного и внутреннего слоев. Внутримембранные частицы – интегральные, полуинтегральные и периферические белки, вкрапленные в бислой фосфолипидов толщиной 7 нм и увеличивающие толщину плазмолеммы до 10 нм. Гликокаликс – надмембранный слой углеводов толщиной 3 нм (в энтероцитах – 50 нм). С белками внутренней поверхности мембраны взаимодействуют компоненты цитоскелета, составляющие плотный кортикальный слой толщиной 100–500 нм.
Функции: барьерная, интегративная (рецепторы для сигнальных молекул, клеточные контакты), транспортная. Транспорт: липиды проникают через плазмолемму свободно, вода – через поры, ионы, глюкоза, аминокислоты – посредством белков-переносчиков, макромолекулы – экзоцитозом (наружу), пиноцитозом (внутрь), частицы, бактерии поглощаются фагоцитозом.
Плазмолемма участвует в простом соединении клеток и по типу замка, формирует сложные контакты (плотные, щелевые, десмосомы). Изолирующие гребешками слияния плотные контакты напоминают стеганое одеяло (на разрезе – застежку «молния»), состоят из белков, встроенных в контактирующие мембраны. Щелевые контакты (нексусы) на сколе мембраны напоминают сито, через поры которого диаметром 2 нм (коннексоны), образованные белками коннексинами, диффундируют ионы. Сцепляющие контакты десмосомы напоминают заклепки, «шляпки» которых (внутриклеточные десмосомные пластинки) образованы структурами подмембранного
12
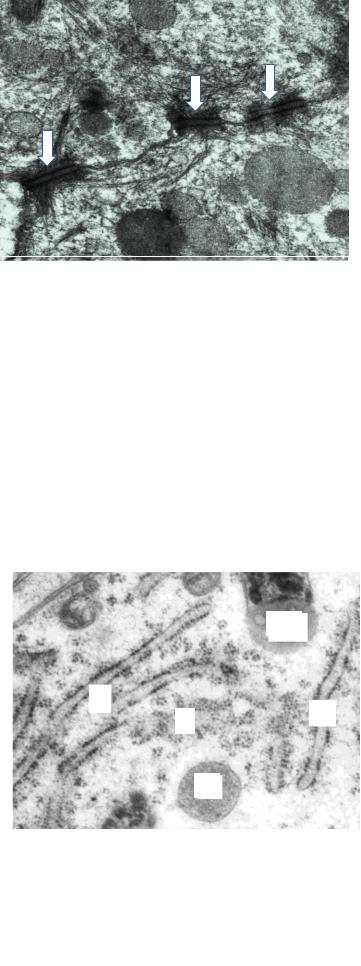
слоя, «стержень» (плотная |
|
|||||
межклеточная линия) – ин- |
|
|||||
тегральными белками (гли- |
|
|||||
копротеидами) |
десмоглеи- |
|
||||
нами (рис. 5). |
|
|
|
|||
Цитоплазма |
объединяет |
|
||||
постоянные |
компоненты |
– |
|
|||
органеллы, |
отвечающие |
за |
|
|||
жизненно |
важные функции |
|
||||
клетки, непостоянные компо- |
|
|||||
ненты – |
цитоплазматиче- |
|
||||
ские включения и матрикс – |
Рис. 5. Десмосомы в эпителиоците вывод- |
|||||
гиалоплазму (цитозоль). |
|
ного протока поднижнечелюстной железы |
||||
|
крысы (обозначены стрелками). Электрон- |
|||||
Органеллы условно клас- |
||||||
ная микрофотография, ув. ×20000 (препарат |
||||||
сифицируют по строению на |
||||||
А.В. Герасимова) |
||||||
мембранные |
(митохондрии, |
|
||||
|
||||||
эндоплазматическая сеть, комплекс Гольджи, лизосомы, пероксисомы) и немембранные (рибосомы, полисомы, клеточный центр, микротрубочки, микрофиламенты). Органеллы общего значения присутствуют во всех клетках, специальные органеллы – в отдельных специализированных клетках.
Эндоплазматическая сеть обнаружена при исследовании в электронном микроскопе внутренней части цитоплазмы фибробласта
американским ученым К. Пор- |
|
|
||||
тером в 1945 г. в виде ограни- |
|
|
||||
ченных мембраной трубочек и |
|
4 |
||||
цистерн с просветом около 20 |
|
|||||
|
|
|||||
нм. Обычно занимает до 10% |
|
|
||||
объема |
цитоплазмы |
(в фиб- |
2 |
|
||
робластах – 50% и более). |
2 |
|||||
1 |
||||||
|
|
|
|
|
||
Американский ученый ру- |
|
|
||||
мынского происхождения Дж. |
|
|
||||
Паладе с помощью более со- |
|
3 |
||||
вершенного |
электронного |
|
|
|||
микроскопа в 1953 г. рассмот- |
Рис. 6. Органеллы в пинеалоците шишко- |
|||||
|
|
|
|
|||
рел на |
поверхности |
цистерн |
видной железы крысы: 1 – свободные рибо- |
|||
гранулярной |
эндоплазматиче- |
сомы, 2 – гранулярная эндоплазматическая |
||||
ской сети тельца – рибосомы |
сеть, 3 – лизосома, 4 – включения липофусцина |
|||||
в телолизосоме. Электронная микрофотогра- |
||||||
(Нобелевская премия, 1974). |
||||||
фия, ув. ×30000 (препарат А.В. Герасимова) |
||||||
13
Рибосомы – электронно-плотные структуры размером 25×20×20 нм, состоят из рРНК и белков. Они обусловливают базофилию цитоплазмы и участвуют в синтезе белка (трансляции), используя в качестве матрицы иРНК. Аминокислоты транспортируют тРНК. Свободные рибосомы и полисомы синтезируют белки опорно-двигательной системы клетки (тубулины, кератин, актин, миозин), белки гиалоплазмы (фибронектин, гельзолин). 75% рибосом удерживает гранулярная эндоплазматическая сеть (ГЭС), синтезирующая экспортные белки, плазмолеммы, лизосомные ферменты. Расширение цистерн свидетельствует об активизации ГЭС (рис. 6).
Рис. 7. Гладкая эндоплазматическая сеть в пинеалоците шишковидной железы мыши (обозначена стрелками). Электронная микро-
фотография, ув. ×30000 (препарат А.В. Герасимова)
1б
1а
Рис. 8. Комплекс Гольджи в секреторном нейроне гипоталамуса крысы: 1а – транс по-
верхность, 1б – цис поверхность. Электронная микрофотография, ув. ×30000 (препарат А.В. Герасимова)
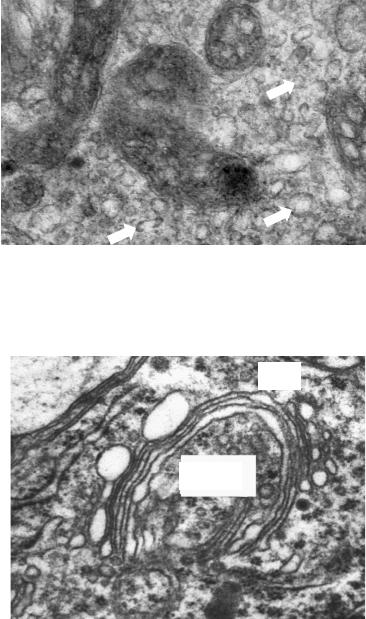
Гладкая эндоплазматическая сеть имеет отно-
шение к синтезу липидов, гликогена, депонированию ионов Са2+, обезвреживанию токсинов, обновлению пероксисом. Ее трубочки лишены рибосом и предопределяют оксифилию цитоплазмы. Расширение трубочек свидетельствует об активизации синтетической деятельности (рис. 7).
Комплекс Гольджи
носит имя первооткрывателя, величайшего нейрогистолога, лауреата Нобелевской премии 1906 г. К. Гольджи (обнаружил в 1898 г. в нервных клетках, импрегнированных серебром). Комплекс объединяет множество диктиосом, состоящих из стопки изогнутых мешочков с расширенными краями. К выпуклой цис поверхности диктиосомы, обращенной к ядру, подходят транспортные пузырьки. Они доставляют
14
белки из ГЭС. Окаймленные пузырьки транспортируют материал плазмолеммы для обновления мембраны мешочков. На вогнутой транс поверхности диктиосомы обнаруживаются лизосомные почки, везикулы Гольджи, транспортирующие лизосомные ферменты («первичные» лизосомы) и секреторные везикулы (рис. 8).
В комплексе Гольджи происходят накопление и сортировка (сегрегация) белков, гликозилирование (образование гликопротеидов).
Лизосомы впервые описал в 1949 г. бельгийский гистохимик С. де Дюв (разделил Нобелевскую премию с Дж. Паладе). Отпочковавшиеся везикулы Гольджи, транспортирующие около 50 лизосомных ферментов (маркерный – кислая фосфатаза), неактивны. Они имеют диаметр 0,2–0,5 мкм, базофильны, с мелкозернистым электронноплотным матриксом, отграниченным от цитозоля мембраной. При закислении цитозоля лизосомная мембрана становится проницаемой, происходит аутолиз клетки – некроз. При слиянии лизосом с фагосомами образуются активные фаголизомы, с органеллами клетки – аутолизосомы. Основная функция лизосом – утилизация структур. Телолизосомы (остаточные, резидуальные тельца) – неактивные, прекратившие переваривать материал лизосомы. Непереваренный материал может структурироваться в пигмент липофусцин, кальцинироваться и выделяться из клетки путем экзоцитоза.
Пероксисомы являются производными гладкой эндоплазматической сети, имеют диаметр 0,3–1,5 мкм, кристаллоидную сердцевину (у крыс), ограничены мембраной. Продолжительность жизни – 5–6 дней. Содержат около 40 различных ферментов, в том числе имеющих отношение к расщеплению Н2О2 (маркерный фермент каталаза), синтезу солей желчных кислот (в гепатоцитах печени).
Митохондрии – самовоспроизводящиеся (почкованием) органеллы размером 0,5×5 мкм, которые окрашиванием кислым фуксином впервые обнаружил немецкий ученый Р. Альтман в 1894 г., а К. Бенда в 1897 г. предложил современное название. Совокупность митохондрий клетки составляет хондриом. Основная функция органелл
– синтез АТФ («болезнь митохондрий» сопровождается разобщением окисления и фосфорилирования, рассеиванием вырабатываемой энергии в виде тепла). Митохондрия имеет рибосомы (миторибосомы), кольцевую ДНК, плотные гранулы кальцификатов в митохондриальном матриксе, митохондриальную оболочку и кристы. Кристы образует внутренняя митохондриальная мембрана, усеянная грибовидными частицами диаметром 8 нм. Наружная мембрана гладкая. Межмембранное пространство составляет 20 нм (рис. 9).
15

Неактивные |
митохон- |
|
|||||
дрии – мелкие, нитевидные, |
|
||||||
с плотно уложенными кри- |
|
||||||
стами. Активные – крупные, |
2 |
||||||
широкие, |
спиралевидные. |
||||||
1 |
|||||||
Погибающие – с просвет- |
|||||||
|
|||||||
ленным матриксом и рас- |
|
||||||
правленными |
|
кристами. |
|
||||
Митохондриальные |
кристы |
|
|||||
обычно не полностью пере- |
|
||||||
гораживают |
полость |
мито- |
|
||||
хондрии. Их ориентация по |
Рис. 9. Митохондрия в пинеалоците шишко- |
||||||
отношению |
к |
длинной |
оси |
||||
видной железы крысы: 1 – кристы, опушенные |
|||||||
митохондрии |
различна |
для |
|||||
грибовидными частицами, 2 – митохондриаль- |
|||||||
разных клеток: перпендику- |
ная оболочка. Электронная микрофотография, |
||||||
лярная (клетки |
печени, |
по- |
ув. ×30000 (препарат А.В. Герасимова) |
||||
|
|||||||
чек), продольная (кардиомиоциты). Кристы могут ветвиться или образовывать пальцевидные
отростки, изгибаться. У простейших, одноклеточных водорослей, в некоторых клетках высших растений и животных (кортикостероциты) выросты внутренней митохондриальной мембраны имеют вид трубок, в гепатоцитах – пластинчатые, в мышечных волокнах мухи – перфорированные, у амебы – волнистые.
Цитоскелет: клеточный центр, микротрубочки, микрофиламенты.
Клеточный центр обна-
ружен в 1875 г. методом ре- |
|
|||||
грессивного |
окрашивания, |
а |
2 |
|||
название |
дал |
Т. Бовери |
в |
|||
1 |
||||||
1895 г. Состоит из двух рас- |
||||||
|
||||||
положенных |
перпендикуляр- |
3 |
||||
но друг к другу центриолей |
||||||
|
||||||
размером 0,1×0,3 мкм (дипло- |
|
|||||
сомы), |
перицентриолярного |
|
||||
матрикса (центросферы) и |
|
|||||
микротрубочек |
астросферы |
|
||||
(рис. 10). |
|
|
|
|
Рис. 10. Клеточный центр в цитоплазме пи- |
|
Каждая |
центриоль объ- |
неалоцита шишковидной железы лесной по- |
||||
единяет |
9 |
периферических |
левки: 1 – диплосома, 2 – центросфера, 3 – |
|||
астросфера. Электронная микрофотография, |
||||||
триплетов |
микротрубочек |
|||||
ув. ×20000 (препарат А.В. Герасимова) |
||||||
(9х3+0). Основная функция –
16
образование микротрубочек, веретена клеточного деления. Микротрубочки состоят из белков тубулинов диаметром 7 нм,
формирующих 13 нитей, расположенных по окружности. Имеют просвет 14 нм и наружный диаметр 28 нм. Определяют форму клетки и перемещают компоненты цитоплазмы, используя механизм скольжения и поперечные «ручки» из белка кинезина. «Ручки» обладают АТФазной активностью.
Микрофиламенты – нити белка актина диаметром 5–7 нм, промежуточные филаменты – нити белка кератина диаметром 7–10 нм, миозин – белковая нить диаметром 10–15 нм с подвижной «головкой», проявляющей АТФ-азную активность. Промежуточные филаменты выполняют опорную функцию. Актино-миозиновые комплексы обеспечивают изменение формы клетки (сократительный аппарат).
Поверхностные специализированные структуры – микровор-
синки и реснички.
Микроворсинки имеет размеры 0,1×1 мкм, продольно расположенные микрофиламенты, выполняющие опорную функцию. Они увеличивают площадь поверхности всасывания веществ (в энтероцитах, нефроцитах).
Реснички размером 0,3×5 мкм с аксонемой и базальным тельцем. Являются подвижными органеллами (25 мерцаний в 1 секунду). Ак-
сонема состоит из 9 перифе- |
|
|
|||||
рических и одного централь- |
1 |
3 |
|||||
ного дублета микротрубочек |
|
4 |
|||||
(9х2+2), |
белковых |
«ручек» |
2 |
||||
|
|||||||
(белок динеин с АТФ-азной |
|
||||||
|
|
||||||
активностью), |
обеспечиваю- |
|
|
||||
щих «скольжение» дублетов |
|
|
|||||
микротрубочек. |
Базальное |
|
|
||||
тельце (кинетосома) анало- |
|
|
|||||
гично по строению центрио- |
|
|
|||||
ли (рис. 11). |
|
|
|
|
|
||
Гиалоплазма – белковая |
Рис. 11. Поверхностные специализирован- |
||||||
коллоидная |
система, |
мат- |
ные структуры: 1 – микроворсинки, 2 – рес- |
||||
рикс, основу |
которого |
со- |
нички, 3 – центральный дублет микротрубочек |
||||
аксонемы, 4 – периферические микротрубочки |
|||||||
ставляют |
микрофиламенты, |
||||||
аксонемы реснички. Электронная микрофото- |
|||||||
объединенные в трехмерную |
графия (по А. Хэму и Д. Кормаку) |
|
|||||
сеть посредством белка фи- |
|
|
|||||
ламина. |
Белок |
гельзолин |
|
|
|||
17
нарушает взаимосвязь филамина с микрофиламентами, гель переходит в золь, что благоприятствует обмену веществ.
Цитоплазматические включения по значимости подразделяют на трофические (резервные накопления питательных веществ – суданофильных, осмиофильных липидных капелек, гликогеновых ШИКпозитивных гранул), секреторные (ШИК-позитивные слизистые везикулы, оксифильные электронно-плотные белковые секреторные везикулы), экскреторные (выделяемые конечные продукты обмена) и пигментные (гемоглобин, липофусцин, меланин, введенный прижизненно (витально) внутривенно краситель трипановый синий).
ОПИСАНИЕ МИКРОПРЕПАРТОВ
1. Кровь человека. Мазок. Окраска азур II – эозином (Прило-
жение, рис. 1). При малом увеличении микроскопа в препарате различимы свободно расположенные мелкие клетки. Это мазок. Численно преобладают в нем безъядерные оксифильно окрашенные эозином в красный цвет округлые клетки – эритроциты. Единичные клетки имеют темное ядро, базофильно окрашенное азуром II в синефиолетовый цвет. Это лейкоциты. Препарат – мазок крови человека. При большом увеличении микроскопа в эритроцитах-дискоцитах в центральной двояковогнутой части различимо просветление. Периферическая часть эритроцитов имеет более интенсивную окраску. В лейкоцитах проявляется полиморфизм клеточных ядер. Чаще других обнаруживаются клетки, размером несколько превосходящие эритроциты с сегментированным ядром и с оксифильно окрашенной в розо- во-фиолетовый цвет мелкозернистой цитоплазмой. Количество сегментов составляет 3–5. Они связаны тончайшими перемычками и имеют околоядерный придаток в виде барабанной палочки – мазок крови женщины. Более мелкие, чем эритроциты, или одинакового с ними диаметра лейкоциты имеют сферическое ядро, окруженное узким ободком базофильной цитоплазмы. Самые крупные лейкоциты содержат крупное почковидное или с несколькими выемками ядро, окруженное широким ободком слабо базофильной цитоплазмы. Просматривая несколько полей зрения, можно обнаружить лейкоциты с палочковидным U-образно или S-образно изогнутым ядром.
Зарисовать на одном рисунке безъядерную клетку эритроцитдискоцит при большом увеличении, на другом – несколько безъядерных эритроцитов и лейкоциты, обозначив полиморфизм их ядер (малый лимфоцит с округлым ядром, окруженным узким ободком цито-
18
плазмы, моноцит с почковидным ядром, окруженным широким ободком слабо базофильной цитоплазмы, нейтрофильные гранулоциты с сегментированным и палочковидным ядром).
2.Однослойный кубический эпителий канальцев почки. Окраска гематоксилином и эозином (Приложение, рис. 2). При ма-
лом увеличении микроскопа в срезе обнаруживаются структуры с округлым просветом – канальцы, характерные для почки кролика. При большом увеличении микроскопа в стенке канальцев различимы расположенные в один ряд клетки. Высота клеток и поперечный размер равнозначны (кубические клетки). Ядра сферической формы, с хроматином, конденсированным в виде мелких глыбок, и ядрышками, базофильно окрашенными гематоксилином в сине-фиолетовый цвет. Оксифильная цитоплазма из-за присутствия в ней белкового цитоскелета и митохондрий, обеспечивающих энергией реабсорбцию веществ, окрашивается кислым красителем эозином в розовый цвет.
Зарисовать при большом увеличении эпителиоциты канальцев почки кубической формы с округлым ядром, сине-фиолетовым ядрышком и глыбками хроматина, оксифильной цитоплазмой, окрашенной в розовый цвет.
3.Однослойный столбчатый эпителий канальцев почки. Окраска гематоксилином и эозином (Приложение, рис. 3). При ма-
лом увеличении микроскопа в срезе обнаруживаются структуры с округлым просветом – канальцы, характерные для почки кролика. При большом увеличении микроскопа в стенке канальцев различимы расположенные в один ряд клетки. Высота клеток превосходит в несколько раз поперечный размер (столбчатые клетки). Ядра овоидной формы вытянуты вдоль оси клеток. Хроматин конденсирован в виде мелких глыбок. Ядрышки и хроматин базофильно окрашиваются гематоксилином в сине-фиолетовый цвет. Оксифильная цитоплазма изза присутствия в ней белкового цитоскелета и большого числа митохондрий, обеспечивающих энергией реабсорбцию веществ, окрашивается кислым красителем эозином в розовый цвет. На апикальной поверхности клеток, обращенной в просвет канальцев, различима поверхностная специализированная структура – щеточная каемка. Она более интенсивно окрашивается эозином и состоит из микроворсинок, увеличивающих площадь поверхности реабсорбции и экскреции веществ в почках.
Зарисовать при большом увеличении столбчатые эпителиоциты канальцев почки с овальным ядром, сине-фиолетовым ядрышком и
19
глыбками хроматина, оксифильной цитоплазмой, окрашенной в розовый цвет.
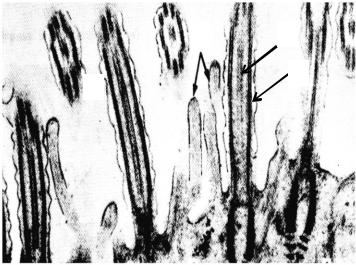
4. Нейрофибриллы в нервных клетках спинного мозга. Импрегнация нитратом серебра (Приложение, рис. 4). При малом уве-
личении микроскопа в срезе органа обнаруживаются центрально расположенное темное вещество, имеющее форму бабочки или буквы «Н», серое вещество, которое окружает более светлое – белое вещество, характерные для спинного мозга собаки, импрегнированного нитратом серебра. В более широких передних рогах серого вещества спинного мозга при большом увеличении микроскопа обнаруживаются группы крупных клеток отростчатой формы – α-мотонейроны. Клетки имеют несколько отростков, в теле нейрона центрально располагается пузырьковидное ядро округлой формы с ядрышком. В перекарионе и отростках выявляется зачерненный серебром цитоскелет в виде тонких нитчатых образований, называемых нейрофибриллами
– органеллы специального значения, характерные только для нервных клеток. В перикарионах нейрофибриллы располагаются в виде плотной сети, окружающей ядро. В отростках нервных клеток нейрофибриллы следуют параллельно друг другу.
Зарисовать при большом увеличении отростчатую нервную клетку с пузырьковидным ядром, ядрышком и зачерненными нейрофибриллами в желто-коричневой цитоплазме. Обозначить нейрофибриллы.
5. Поперечнополосатая мышечная ткань языка. Окраска же-
лезным гематоксилином (Приложение, рис. 5). При малом увеличении микроскопа в препарате отыскать продольно и поперечно разрезанные толстые темно-синего цвета волокна, характерные для мышечной ткани языка кролика, окрашенной железным гематоксилином. Мышечные волокна собраны в пучки и разделены светлыми прослойками. При большом увеличении микроскопа в продольно разрезанных мышечных волокнах различимы узкие поперечные темные и светлые диски, состоящие из толстых миозиновых миофиламентов и тонких актиновых миофиламентов. Миофиламенты организуются в поперечнополосатые миофибриллы – органеллы специального значения, составляющие сократительный аппарат. Миофибриллы простираются вдоль центральной части волокон без перерыва на всем их протяжении. Снаружи от миофибрилл в общей цитоплазме располагаются многочисленные ядра овоидной формы с умеренно конденсированным хроматином, характерные для миосимпластов, производных клеток миобластов, образующих многоядерные струк-
20