

Книги по МРТ КТ на английском языке / PLUM AND POSNER S DIAGNOSIS OF STUPOR AND COM
.pdf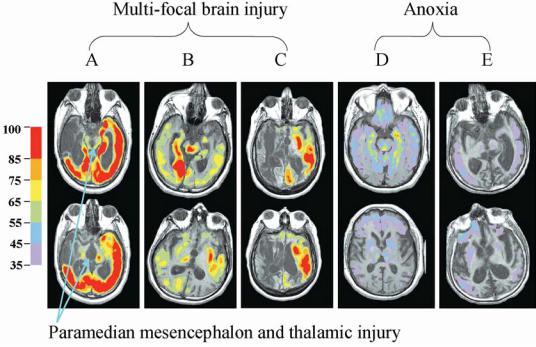
374 |
Plum and Posner’s Diagnosis of Stupor and Coma |
||
The very low overall resting cerebral met- |
arousal systems.145,146 Even incomplete in- |
||
abolic rates in MCS patients generally in- |
juries to these networks may produce unique |
||
clude the posterior and ventroanterior cingu- |
deficits in maintaining adequate cerebral ac- |
||
late regions associated by Raichle with self- |
tivation and patterns of brain dynamics neces- |
||
awareness. This may account for the failure to |
sary to establish, maintain, and complete com- |
||
engage functional network activation with pre- |
plex behaviors. |
||
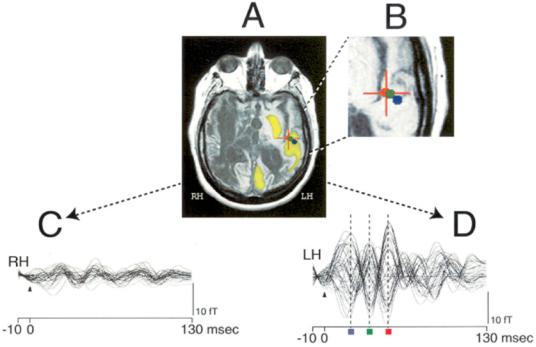
sentation of time-reversed narratives (Figure |
|
||
9–10). Specifically, a lack of a metabolically ex- |
|
||
pensive ongoing self and environmental mon- |
The Potential Role of Regionally |
||
itoring process may leave the MCS brain stim- |
|||
Selective Injuries Producing |
|||
ulus bound and limited to activations provoked |
|||
Widespread Effects on |
|||
by extremely salient events. This interpretation |
|||
is supported by direct comparisons of changes |
Brain Function |
||
in cerebral metabolism, functional MRI signal |
|
||
activation, and neuronal activity that indicate |
At least three different mechanisms may lead |
||
a linear correlation of these measures.142,143 |
to marked alteration of integrative brain ac- |
||
The dissociation of low resting cerebral me- |
tivity following relatively focal or regionally |
||
tabolism and recruitable cerebral networks in |
restricted brain lesions: (1) a form of passive |
||
MCS invites speculation that patients who re- |
inhibition of a brain area following deaffer- |
||
main near the border of emergence from MCS |
entation of remote but strongly connected ar- |
||
(see red line in Figure 9–1) may show fluctu- |
eas, (2) active inhibitory phenomena resulting |
||
ation of recruitment of these large-scale net- |
from altered connectivity and neuronal func- |
||
works under varying internal conditions of |
tion following injury, and (3) persistent or par- |
||
arousal and appearance of environmentally sa- |
oxysmal functional activity producing excess |
||
lient stimuli, leading to the occasional surpris- |
excitation of distributed neuronal networks.121 |
||
ingly high level of response. |
Whether such processes underlie partially re- |
||
A further consideration is whether injuries |
versible impairment of cognitive function in |
||
incurred by compression of the thalamus and |
severely disabled patients is unknown. It is |
||
brainstem during acute herniation may un- |
likely, however, that transient changes in dis- |
||
derlie the chronically low metabolic rates in |
tributed network function underlie the wide |
||
patients remaining in MCS despite connected |
fluctuations in cognitive performance in some |
||
and recruitable cerebral networks (both MCS |
MCS patients and patients who emerge from |
||
patients studied121 had herniated with mid- |
MCS. These phenomena are well known but |
||
brain signs of third nerve palsies during the |
not frequently described in the medical liter- |
||
acute phase of their injuries). As discussed in |
ature.91,127 We briefly discuss potentially rel- |
||
Chapter 1, the paramedian mesencephalon and |
evant sources of variations of brain dynamics |
||
thalamus contain several interconnected brain |
within the wakeful state of the injured brain. |
||
systems that interact closely with the brain- |
A relatively common finding following focal |
||
stem arousal systems. Although these struc- |
ischemia or traumatic brain injury is a reduc- |
||
tures were originally identified as the primary |
tion in cerebral metabolism in brain regions |
||
arousal systems, the thalamic intralaminar nu- |
remote from the site of injury. This transsy- |
||
clei (ILN) (and paralaminar regions of the |
naptic (or ‘‘crossed’’) down-regulation of dis- |
||
thalamus rich in neurons that preferentially |
tant neuronal populations results from the loss |
||
project to layer I of the cerebral cortex), the |
of excitatory inputs from the damaged re- |
||
mesencephalic reticular formation (MRF), |
gions.147 The clinical significance of these |
||
and their connections with the thalamic retic- |
changes is unclear, although electrophysiologic |
||
ular nucleus appear to play a key role linking |
correlates have been identified. A recent study |
||
arousal states to the control of moment-to- |
by Gold and Lauritzen148 showed that al- |
||
moment intention or attentional gating (re- |
though changes in blood flow may be modest |
||
viewed in 144). These structures are well posi- |
in remote cortical regions, the transsynaptic |
||
tioned to control interactions of the cerebral |
down-regulation produces dramatic decreases |
||
cortex, basal ganglia, and thalamus through |
in neuronal firing rates (e.g., a neuronal firing |
||
their patterns of innervation within the cortex |
rate decreased by 80% with only a 20% re- |
||
as well as rich innervation from the brainstem |
duction in regional blood flow). Thus, stable |
||