

18.Репарация как механизм поддержания генетического гомеостаза. Виды репарации. Мутации, связанные с нарушением репарации и их роль в патологии.
Для поддержания главных характеристик клетки или организма на протяжении их жизни, а также в ряду поколений наследственный материал должен отличаться устойчивостью к внешним воздействиям или должны существовать механизмы коррекции возникающих в нем изменений. В живой природе используются оба фактора. Третьим фактором является точность копирования нуклеотидных последовательностей материнской ДНК в процессе ее репликации.
ДНК-геликаза расплетает двойную спираль ДНК, разделяя ее полинуклеотидные цепи; дестабилизирующие белки выпрямляют участок цепи ДНК; ДНК-топоизомераза разрывает фосфодиэфирную связь в одной из полинуглеотидных цепей ДНК, снимая напряжение, вызываемое расплетенисм спирали и расхождением цепей в репликационной вилке; РНК-праймаза синтезирует РНК-затравки для дочерней цепи и для каждого фрагмента Оказаки; ДНК-полимераза осуществляет непрерывный синтез лидирующей цепи и синтез фрагментов Оказаки отстающей цепи; ДНК-лигаза сшивает фрагменты Оказаки после удаления РНК-затравки.
По реакционной способности молекулы ДНК относятся к категории химически инертных веществ. Известно, что роль вещества наследственности может выполнять не только ДНК, но и РНК (некоторые вирусы). Считают, что выбор в пользу ДНК обусловлен ее более низкой по сравнению с РНК реакционной способностью.
В поддержании высокой точности репликации важная роль принадлежит прежде всего ферменту ДНК-полимеразе. Этот фермент осуществляет отбор необходимых нуклеотидов из числа имеющихся в ядерном соке нуклеозидтрифосфатов (АТФ, ТТФ, ГТФ, ЦТФ), точное присоединение их к матричной цепи ДНК и включение в растущую дочернюю цепь. Частота включения неправильных нуклеотидов на этой стадии составляет 1·10-5 пар оснований.
Такие ошибки в работе ДНК-полимеразы связаны с возникновением измененных форм азотистых оснований, которые образуют «незаконные» пары с основаниями материнской цепи. Например, измененная форма цитозина вместо гуанина связывается водородными связями с аденином. В результате в растущую цепь ДНК включается ошибочный нуклеотид. Быстрый переход измененной формы такого основания в обычную нарушает его связывание с матрицей, появляется неспаренный 3'-ОН-конец растущей цепи ДНК. В этой ситуации включается механизм самокоррекции, осуществляемый ДНК-полимеразой (или тесно связанным с ней ферментом — редактирующей эндонуклеазой). Самокоррекция заключается в отщеплении ошибочно включенного в цепь ДНК нуклеотида, не спаренного с матрицей. Следствием самокоррекции является снижение частоты ошибок в 10 раз (с 10-5 до 10-6).
Несмотря на эффективность самокоррекции, в ходе репликации после удвоения ДНК в ней обнаруживаются ошибки. Особенно часто это наблюдается при нарушении концентрации четырех нуклеозидтрифосфатов в окружающем субстрате. Значительная часть изменений возникает также в молекулах ДНК в результате спонтанно происходящих процессов, связанных с потерей пуриновых оснований — аденина и гуанина (апуринизацией) — или дезаминированием цитозина, который превращается в урацил. Частота последних изменений достигает 100 на 1 геном/сут.
Содержащиеся в ДНК основания могут изменяться под влиянием реакционноспособных соединений, нарушающих их нормальное спаривание, а также под действием ультрафиолетового излучения, которое может вызвать образование ковалентной связи между двумя соседними остатками тимина в ДНК (димеры тимина). Названные изменения в очередном цикле репликации должны привести либо к выпадению пар оснований в дочерней ДНК, либо к замене одних пар другими. Указанные изменения действительно сопровождают каждый цикл репликации ДНК, однако их частота значительно меньше, чем должна была бы быть. Это объясняется тем, что большинство изменений такого рода устраняется благодаря действию механизма репарации (молекулярного восстановления) исходной нуклеотидной последовательности ДНК.
Механизм репарации основан на наличии в молекуле ДНК двух комплементарных цепей. Искажение последовательности нуклеотидов в одной из них обнаруживается специфическими ферментами. Затем соответствующий участок удаляется и замещается новым, синтезированным на второй комплементарной цепи ДНК. Такую репарацию называют эксцизионной, т.е. с «вырезанием». Она осуществляется до очередного цикла репликации, поэтому ее называют также дорепликативной.
Восстановление исходной структуры ДНК требует участия ряда ферментов. Важным моментом в запуске механизма репарации является обнаружение ошибки в структуре ДНК. Нередко такие ошибки возникают во вновь синтезированной цепи в процессе репликации. Ферменты репарации должны обнаружить именно эту цепь. У многих видов живых организмов вновь синтезированная цепь ДНК отличается от материнской степенью метилирования ее азотистых оснований, которое отстает от синтеза. Репарации при этом подвергается неметилированная цепь. Объектом узнавания ферментами репарации могут также служить разрывы в цепи ДНК. У высших организмов, где синтез ДНК происходит не непрерывно, а отдельными репликонами, вновь синтезируемая цепь ДНК имеет разрывы, что делает возможным ее узнавание.
Восстановление структуры ДНК при утрате пуриновых оснований одной из ее цепей предполагает обнаружение дефекта с помощью фермента эндонуклеазы, которая разрывает фосфоэфирную связь в месте повреждения цепи. Затем измененный участок с несколькими примыкающими к нему нуклеотидами удаляется ферментом экзонуклеазой, а на его месте в соответствии с порядком оснований комплементарной цепи образуется правильная нуклеотидная последовательность
При изменении одного из оснований в цепи ДНК в восстановлении исходной структуры принимают участие ферменты ДНК-гликозилазы числом около 20. Они специфически узнают повреждения, обусловленные дезаминированием, алкилированием и другими структурными преобразованиями оснований. Такие модифицированные основания удаляются. Возникают участки, лишенные оснований, которые репарируются, как при утрате пуринов. Если восстановление нормальной структуры не осуществляется, например в случае дезаминирования азотистых оснований, происходит замена одних пар комплементарных оснований другими —пара Ц—Г может заменяться парой Т—А и т.п.
Образование в полинуклеотидных цепях под действием УФ-лучей тиминовых димеров (Т—Т) требует участия ферментов, узнающих не отдельные измененные основания, а более протяженные повреждения структуры ДНК. Репаративный процесс в этом случае также связан с удалением участка, несущего димер, и восстановлением нормальной последовательности нуклеотидов путем синтеза на комплементарной цепи ДНК.
В том случае, когда система эксцизионной репарации не исправляет изменения, возникшего в одной цепи ДНК, в ходе репликации происходит фиксация этого изменения и оно становится достоянием обеих цепей ДНК. Это приводит к замене одной пары комплементарных нуклеотидов на другую либо к появлению разрывов (брешей) во вновь синтезированной цепи против измененных участков. Восстановление нормальной структуры ДНК при этом может произойти и после репликации.
Пострепликативная репарация осуществляется путем рекомбинации (обмена фрагментами) между двумя вновь образованными двойными спиралями ДНК. Примером такой пострепликативной репарации может служить восстановление нормальной структуры ДНК при возникновении тиминовых димеров (Т—Т), когда они не устраняются самопроизвольно под действием видимого света (световая репарация) или в ходе дорепликативной эксцизионной репарации.
Ковалентные связи, возникающие между рядом стоящими остатками тимина, делают их не способными к связыванию с комплементарными нуклеотидами. В результате во вновь синтезируемой цепи ДНК появляются разрывы (бреши), узнаваемые ферментами репарации. Восстановление целостности новой полинуклеотидной цепи одной из дочерних ДНК осуществляется благодаря рекомбинации с соответствующей ей нормальной материнской цепью другой дочерней ДНК. Образовавшийся в материнской цепи пробел заполняется затем путем синтеза на комплементарной ей полинуклеотидной цепи. Проявлением такой пострепликативной репарации, осуществляемой путем рекомбинации между цепями двух дочерних молекул ДНК, можно считать нередко наблюдаемый обмен материалом между сестринскими хроматидами.
В ходе дорепликативной и пострепликативной репарации восстанавливается большая часть повреждений структуры ДНК. Однако, если в наследственном материале клетки возникает слишком много повреждений и часть из них не ликвидируется, включается система индуцируемых (побуждаемых) ферментов репарации (SOS-система). Эти ферменты заполняют бреши, восстанавливая целостность синтезируемых полинуклеотидных цепей без точного соблюдения принципа комплементарности. Вот почему иногда сами процессы репарации могут служить источником стойких изменений в структуре ДНК (мутаций). Названная реакция также относится к SOS-системе.
Если в клетке, несмотря на осуществляемую репарацию, количество повреждений структуры ДНК остается высоким, в ней блокируются процессы репликации ДНК. Такая клетка не делится, а значит, не передает возникших изменений потомству.
Вызываемая повреждениями ДНК остановка клеточного цикла в сочетании с невозможностью молекулярной репарации измененного наследственного материала может с участием белка, синтез которого контролируется геном р53, приводить к активации процесса самоликвидации (апотпоз) дефектной клетки с целью устранения ее из организма.
Таким образом, обширный набор различных ферментов репарации осуществляет непрерывный «осмотр» ДНК, удаляя из нее поврежденные участки и способствуя поддержанию стабильности наследственного материала. Совместное действие ферментов репликации (ДНК-полимераза и редактирующая эндонуклеаза) и ферментов репарации обеспечивает достаточно низкую частоту ошибок в молекулах ДНК, которая поддерживается на уровне 1 · 10-9 пар измененных нуклеотидов на геном. При размере генома человека 3 · 109 нуклеотидных пар это означает появление около 3 ошибок на реплицирующийся геном. Вместе с тем даже этот уровень достаточен для образования за время существования жизни на Земле значительного генетического разнообразия в виде генных мутаций.
№ 19. Ген – участок молекулы ДНК, который несет информацию о структуре полипептидной цепи или макромолекулы. Свойства: 1.Ген – функциональная единица наследственной информации. 2.Ген занимает определенный участок в хромосоме – локус. Гены располагаются в хромосоме в линейном порядке и образуют группу сцепления. Число групп сцепления соответствует гаплоидному набору хромосом. 3. Гены не меняются. Ошибки исправляются репарационными механизмами. 4. Гены способны к мутациям. 5. Полиаллелизм– присутствие в генофонде вида одновременно различных аллелей гена. 6. Каждый ген отвечает за развитие определённого признака или признаков
Генетический код - способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов. Свойства: 1. Триплетность — единица кода - сочетание трёх нуклеотидов (триплет, или кодон). 2. Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно. 3. Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте. 4. Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов. 5. Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии).
Структура РНК. Одна полинуклеотидная цепь, состоящая из 4 пар разновидностей нуклеотидов, содержащих рибозу, фосфат и одно из 4-х азотистых оснований – аденин, гуанин, цитозин, урацил. Азотистые основания в составе РНК могут образовывать водородные связи между Ц и Г, А и У. Есть гидроксильная группа в 2' положении рибозы, которая позволяет молекуле РНК существовать в А-конформации. Виды РНК:
мРНК (иРНК) — РНК, содержащая информацию о первичной структуре (аминокислотной последовательности) белков. мРНК синтезируется на основе ДНК в ходе транскрипции, используется в ходе трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов. Транскрипцией называют процесс копирования генетической информации с ДНК на РНК, в частности на мРНК. Транскрипция осуществляется ферментом РНК-полимеразой, строящей, согласно принципу комплементарности, копию участка ДНК на основании одной из цепей двойной спирали. Сначала возникает про-и-РНК. Это объясняется тем, что у эукариот и-РНК образуется в результате процессинга (созревания). Ген имеет прерывистую структуру. Кодирующие участки – экзоны и некодирующие – интроны. Ген у эукариоических организмов имеет экзонно-интронную структуру. Длина интрона больше длины экзона. В процессе процессинга интроны «вырезаются» - сплайсинг. Иногда интроны одного гена являются экзонами другого, тогда сплайсинг невозможен.
тРНК - трансляционный посредник, доставляет амк к месту сборки полипептидов. тРНК синтезируются на определенных последовательностях ДНК. Состоит из небольшого числа нуклеотидов – 75-95. В форме листа клевера, к концу акцепторного «стебля» присоединяется амк. Противоположный конец тРНК называется антикодоном и несет информацию о триплете, соответствующем данной аминокислоте. 10-15% РНК в клетке.
рРНК образуют сложные комплексы рибосом, на которых происходит трансляция. Белки образуют каркас рибосом и играют роль ферментов. Основной функцией рРНК является осуществление процесса трансляции — считывания информации с мРНК при помощи тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами. Рибосомные субчастицы содержат по одной молекуле рРНК большой длины. рРНК синтезируется в ядрышке.
Поцессинг и сплайсинг способны объединять структуры, удаленные друг от друга, в один ген, поэтому они имеют огромное эволюционное значение. Подобные процессы упрощают видообразование. Например, на границе между 2мя генами фермента ДНК-полимеразы находится интрон; фермент состоит из 2 доменов, которые образуют 2 независимые компактные частицы, связанные полипептидным мостиком. Когда-то домены были раздельными генами, а затем – сблизились. Нарушения подобной структуры гена приводит к генным болезням. Нарушение строения интрона фенотипически незаметно, нарушение в экзонной последовательности приводят к мутации (мутации глобиновых генов).
Роль РНК в процессе реализации наследственной информации. ДНК – макромолекула, она не может выходить в цитоплазму из ядра и передавать информацию. РНК переводит наследственную информацию из ДНК в рабочую форму – синтезируют белки. Различные виды РНК принимают участие в реализации наследственной информации через биосинтез белка.
№ 20. Рибосомный цикл синтеза белка. Перенос информации с иРНК на белок во время его синтеза называется трансляцией. Собранные в полисомы рибосомы двигаются по иРНК; движение происходит последовательно, по триплетам. В месте контакта рибосомы с иРНК работает фермент, собирающий белок из аминокислот, доставляемых к рибосомам тРНК. При этом происходит сравнение кодона иРНК с антикодоном тРНК; если они комплементарны, фермент (синтетаза) «сшивает» аминокислоты, а рибосома продвигается вперед на один кодон. Синтез одной молекулы белка обычно идет 1–2 мин (один шаг занимает 0,2 с)
Фаза инициации начинается со слияния двух субчастиц рибосомы на мРНК, присоединения к мРНК первой тРНК, а также определения рамки считывания информации с мРНК. На 5-м конце любой мРНК находится участок комплементарный рРНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается стартовый кодон АУГ, шифрующий аминокислоты метионин. При этом только тРНК, несущая метионин (у прокариот формилметионин), способна соединиться с малой субчастицей после чего происходит объединение большой и малой субчастиц рибосомы. Процессы инициации катализируются особыми белками - факторами инициации, которые подвижно связаны с малой субчастицей рибосомы. По завершении фазы инициации белки отделяются от рибосомы.
Фаза элонгации представляет собой циклически повторяющиеся события при которых происходит удлинение пептида. тРНК, несущая аминокислоту, при соединении ее антикодона с кодоном мРНК становится в положение 1 большой субчастицы. Между аминокислотой из пептидной цепочки и аминокислотой, соединенной с тРНК, образуется пептидная связь. В результате предыдущая теряет связь со своей тРНК и присоединяется к тРНК, находящейся в положении 1. Далее рибосома продвигается вдоль мРНК и тРНК с цепочкой аминокислот становится в положение 2. Такая последовательность событий повторяется до тех пор, пока в положение 1 рибосомы не поступит кодон-терминатор, для которого не существует соответствующей тРНК.
Фаза терминации связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ, УГА). При этом к последней аминокислоте в пептидной цепи присоединяется вода и ее карбоксильный конец отделяется от тРНК, а рибосома распадается на две субчастицы. Синтез пептида происходит не одной рибосомой, а несколькими тысячами, которые образуют комплекс - полисому.
Посттрансляционные преобразования белков. После завершения трансляции и высвобождения белка из рибосомы аминокислоты в составе полипептидной цепи подвергаются разнообразным химическим модификациям. Примерами посттрансляционной модификации являются:
- присоединение различных функциональных групп (ацетил-,метил-ифосфатныхгрупп);
- присоединение липидовиуглеводородов;
- изменение стандартных аминокислот на нестандартные (образование цитруллина);
- образование структурных изменений (образование дисульфидных мостиков между цистеинами);
- удаление части белка как в начале (сигнальная последовательность), так и в отдельных случаях в середине (инсулин);
- добавление небольших белков, которые влияют на деградацию белков.
При этом тип модификации может быть как универсальным, так и специфическим для данного белка. В то же время один и тот же белок может подвергаться многочисленным модификациям. Так, гистоны(белки, входящие в составхроматинау эукариот) в разных условиях могут подвергаться до 150 различных модификаций.
№ 21 Взаимосвязь между геном и признаком. Связь между геном и признаком (продуктом) была открыта при изучении брожения в безвоздушной среде в 1902 г Гарродом. Он изучал родословные больных алкаптонурией, пришел к выводу, что болезнь - результат нарушения обмена азота, при этом вместо мочевины образуется темное вещество. При содействии Бэтса в 1908 году высказано предположение, что болезнь возникает у рецессивных гомозигот, у которых не хватает какой-то ферментативной реакции, что приводит к накоплению и выведению субстрата, который в норме должен был расщепиться. В крови людей содержится гомогентизиновая кислота, но в норме она расщепляется оксидазой гомогентизиновой кислоты до малеинацетата, затем до воды и углекислого газа. У больных нет оксидазы, поэтому происходит накопление кислоты и вывод ее с мочой. Так же наследуется альбинизм, хотя встречается гораздо чаще. При этом заболевании отсутствует фермент, осуществляющий превращение тирозина в меланин.
Гипотеза «один ген – один фермент». В 1940 год - Бидл и Татум предложили гипотезу: 1 ген – 1 фермент. Эта гипотеза сыграла важную роль – ученые стали рассматривать конечные продукты. Оказалось, что гипотеза имеет ограничения, т.к. все ферменты – белки, но не все белки – ферменты. Как правило, белки являются олигомерами – т.е. существуют в четвертичной структуре, в образовании которой принимают участие разные пептидные цепи. Например, гемоглобин взрослого человека включает четыре глобиновых цепи — 2α и 2β, кодируемые разными генами. Поэтому формула, отражающая связь между геном и признаком, была несколько преобразована: «Один ген – одна полипептидная цепь».
№ 22. Ген как единица изменчивости. Ген - наименьший участок хромосомы, способный претерпеть мутацию. Внезапные спонтанные изменения фенотипа, которые нельзя связать с обычными генетическими явлениями, можно объяснить только изменениями в структуре отдельных генов. Такое изменение последовательности оснований в данном гене воспроизводится при транскрипции в структуре мРНК и приводит к изменению последовательности аминокислот в полипептидной цепи, образующейся в результате трансляции на рибосомах.
Генные мутации связаны с изменением внутренней структуры генов, что превращает одни аллели в другие. Можно выделить несколько типов генных мутаций на молекулярном уровне:
- замена пар нуклеотидов. Замена пуринового основания на др. пуриновое, или одного пиримидинового на др. пиримидиновое – транзиция. Замена пуринового основания на пиримидиновое и наоборот – трансверсия. При замене нуклеотидов в структурных генах происходит изменение смысла гена – возникают миссенс-мутации. При этом одна аминокислота в полипептиде замещается другой. Фенотипическое проявление мутации зависит от положения аминокислоты в полипептиде. При замене последовательности ЦТЦ на ЦАЦ возникает серповидно-клеточная анемия. Образуется новый полипептид и гемоглобин имеет совсем другие свойства. Некоторые миссенс-мутации приводят к возникновению фермента, обладающего высокой активностью в одних условиях и средней в других условиях. Т.к. генетический код вырожден, то при замене триплетов, кодирующий одну и ту же аминокислоту, мутации не проявляются. Другой вид мутаций – нонсенс – мутации - при замене одного нуклеотида другим образуются бессмысленные триплеты. Синтез полипептида прекращается и белок имеет совсем иные свойства. УАГ. УАА. УГА. бессмысленные триплеты.
- делеция или вставка одного или нескольких нуклеотидов ведут за собой утрату или вставку одной или нескольких аминокислот в полипептиде. Эффекта может не быть. Если происходят делеция или вставка 1 нуклеотида (или другого числа нуклеотидов не кратного 3), наблюдается сдвиг рамки считывания, при этом нарушается структура полипептида. В организме происходит большое количество мутаций. Они затрагивают интеллект, поведение, метаболические признаки и т.д. мутации, изменяющие видимые морфологические признаки – видимые (мутация альбинизма).
- вставка нуклеотида.
- перестановка (инверсия) участка гена.
Последствия генных мутаций. Генные мутации, возникающие в гаметах или в будущих половых клетках, передаются всем клеткам потомков и могут влиять на дальнейшую судьбу популяции. Соматические генные мутации, происходящие в организме, наследуются только теми клетками, которые образуются из мутантной клетки путем митоза. Они могут оказать воздействие на тот организм, в котором они возникли, но со смертью особи исчезают из генофонда популяции. Мутации, нарушающие жизнь – летальные, полулетальные и сублетальные. Летальные – гибель зиготы или развившегося организма на определенной стадии эмбриогенеза – выкидыши. Полулетальные и сублетальные ослабляют жизнеспособность организма или отдельных клеток (например, брахидактилия – гомозиготы погибают).
№ 23. Хромосома, ее химический состав. Состоит из ДНК и белков, которые образуют нуклеопротеиновый комплекс—хроматин. Белки составляют значительную часть вещества хромосом. На их долю приходится около 65% массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки. Гистоны представлены пятью фракциями: HI, Н2А, Н2В, НЗ, Н4. Они прочно соединяются с молекулами ДНК, препятствуя считыванию заключенной в ней биологической информации; выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах. Число фракций негистоновых белков превышает 100. Помимо ДНК и белков в составе хромосом обнаруживаются также РНК, липиды, полисахариды, ионы металлов.
Структурная организация хроматина:
Нуклеосомный уровень. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных гистонов: Н2А, Н2В, НЗ, Н4. Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов). Молекула ДНК спирально накручивается на коры. При этом в контакте с каждым кором оказывается участок ДНК, состоящий из 146 пар нуклеотидов (п.н.). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают от 15 до 100 п.н. (в среднем 60 п.н.) в зависимости от типа клетки. Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Благодаря такой организации в основе структуры хроматина лежит нить, представляющая собой цепочку повторяющихся единиц — нуклеосом. Диаметр 10—11 нм.
Нуклеомерный. Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном HI, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная по типу соленоида - хроматиновая фибрилла, называемая также элементарной. Диаметр 20—30 нм.
Петельный. Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы. Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 п. н. Диаметр 100—200 нм.
Эухроматин – меньшая плотность упаковки и высокая активность.
Гетерохроматин - компактная организация и генетическая инертность. В его пределах транскрипции биологической информации не происходит.
Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина (1:10000). Отдельные хромосомы становятся хорошо различимы. Диаметр 500—600 нм.
Морфология хромосом. Хромосома состоит из 2 хроматид, соединенных центромерой (кинетохором). Хромосомы бывают:
- метацентрические = равноплечие (центромера посередине);
- субметацентрические = неравноплечие (с центромерой, сдвинутой к одному из концов);
- акроцентрические (с центромерой, расположенной практически на конце хромосомы).
№ 24. Хромосомные мутации (хромосомные аберрации) – структурные перестройки, затрагивающие одну или несколько хромосом, все они связаны с потерей, либо с добавлением участка хромосомы. Аномалии по крупным хромосомам происходят значительно реже, чем по мелким. Самая маленькая хромосома – 21, нарушения ее строения встречаются чаще всего. Нехватка генетического материала переносится хуже, чем избыток. Если много эухроматина – нежизнеспособность ребенка, если преобладает гетерохроматин – тяжелые патологии (8,13,18,21,х хромосомы).
Дилеция - нарушение кроссинговера, при котором отдельные участки хромосом выпадают.
Р – длинное плечо, Q – короткое.
46,хх,5р – дилеция плеча 5 хромосомы. Синдром Кошачий крик. Широко расставленные глаза, физическое недоразвитие. Множественные пороки развития, недоразвита гортань – специфический крик.
Дупликация – удвоение части генетического материала.
Транслокация – обмен участками хромосом:
Реципрокные (обмен участками между негомологичными хромосомами). 46,ху,t(9,22) – миелолейкоз (рак крови).
Нереципроксные (между 2мя гомологичными хромосомами). Может не проявляться.
Робертсоновские: возникают при нарушениях деления акроцентрических хромосом. Разрыв по центромере, короткие части дегенерируют, длинные срастаются часто по 15 хромосоме (46,хх,15t – рак крови).
Инверсия – поворот участка на 180°. 46,хх,inv16. Может являться причиной нарушения процесса конъюгации (образование бивалента) во время мейоза, действуя как “ингибитор кроссинговера”, а в некоторых случаях приводя к формированию нежизнеспособных гамет.
Кольцевые хромосомы могут возникать по 16й и 18й хромосомам, терминальные концы разрываются и склеиваются. 46,хх,r18 – слабоумие, аномалии лица.
Фрагильность (ломкость) по Х-хромосоме. Понижение интеллекта.
Изохромосомы – разделение хромосомы неправильным путем. Чем больше возраст отца, тем, чаще встречается подобное нарушение.
Роль хромосомных мутаций. Приводят к физическим и психическим нарушениям развития. Дупликации играют особую роль в эволюции. Это связано с тем, что они увеличивают количество генетического материала и тем самым открывают возможность возникновения новых генов с новыми свойствами. Некоторые копии оказались полезными, и естественный отбор поддерживал их в популяциях. Были нейтральные копии, присутствие которых никак не сказывалось на приспособленности их носителей, они становились резервом эволюции. Со временем они могли приобретать новые функции и становиться все более и более уникальными. Пример - многочисленное и разнообразное семейство генов глобинов млекопитающих. Анализ последовательности нуклеотидов в этих генах показывается, что все они произошли в результате серии последовательных удвоений одного-единственного гена. За каждым удвоением следовало накопление случайных мутаций и постепенное изменение их функций, синтезируемых ими белков → усложнение организмов.
№ 25. Геном, кариотип. Геном – совокупность генов, характерных для гаплоидного набора хромосом данного вида. При оплодотворении геномы родителей объединяются и образуют клеточный генотип зиготы. Кариотип – диплоидный набор хромосом, характеризующийся совокупностью признаков: число, форма, размер, особенности строения хромосом. Постоянство кариотипа поддерживается механизмам и митоза и мейоза. Каждый вид организмов обладает характерным и постоянным набором хромосом. Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где значение п различно у разных видов. Являясь видовой характеристикой организмов, кариотип может отличаться у отдельных особей некоторыми частными особенностями. Например, у представителей разного пола, имеются в основном одинаковые пары хромосом (аутосомы), но их кариотипы отличаются по одной паре хромосом (гетерохромосомы, или половые хромосомы). Чаще различия касаются строения половых хромосом, обозначаемых разными буквами —X и Y (XX или XY). Идиограмма - схематическое изображение гаплоидного набора хромосом организма, которые располагают в ряд в соответствии с их размерами.
Характеристика кариотипа человека в норме. 46,XX (женский) и 46,XY (мужской).
Классификация хромосом человека по размеру и расположению центромеры: Гр. хромосом Номер по кариотипу Характеристика хромосом А(I) 1,2,3 1 и 3 почти метацентрические и 2—крупная субметацентрическая В(II) 4,5 крупные субакроцентрические С(III) 6—12 средние субметацентрические A(lV) 13—15 средние акроцентрические E(V) 16-18 мелкие субметацентрические F(VI) 19—20 самые мелкие мегацентрические G(VII) 21—22 самые мелкие акроцентрические Х-хромосома (относится к III группе) 23 средняя почти метацентрическая Y-хромосома 23 мелкая акроцентрическая
№ 26. Геном - совокупность генов, характерных для гаплоидного набора хромосом данного вида.
Геном состоит из 3-х с лишним миллиардов нуклеотидов. У человека 35 тыс. генов, хранящих информацию обо всех частях нашего тела и их функции.
Структурные гены содержат информацию о структуре полипептидной цепи (структурных белках).
Регуляторные гены определяют место, время, длительность включения структурных генов.
Регуляция экспрессии генов у прокариот. Французские микробиологи Ф. Жакоб и Ж. Моно (1961) - оперонная модель регуляции транскрипции. Оперон — это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые осуществляют последовательные этапы биосинтеза какого-либо метаболита.
Жакоб и Моно разделили гены регуляторной системы на два типа - гены-регуляторы и гены-операторы. Они предложил, что есть участок молекулы ДНК с группой структурных генов. К этой группе примыкает участок в 200пар нуклеотидов – промотор (участок примыкания ДНК зависимой РНК-полимеразы). К этому участку примыкает ген-оператор. Название всей системы – оперон. Регуляция осуществляется регуляторным геном. В итоге белок-репрессор взаимодействует с геном-оператором, и оперон начинает работать. Субстрат взаимодействует с геном регуляторами, оперон блокируется. Принцип обратной связи. Экспрессия оперона включается как единое целое.
 оперон
оперон
Ген-регулятор (ГР) Ген-оператор(ГО)|промотор|Ген структурный(ГС1) |ГС2 | ГС3





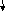 м-РНК
м-РНК1 м-РНК2 м-РНК3
м-РНК
м-РНК1 м-РНК2 м-РНК3



б елок-репрессор
Е1 Е2 Е3
(белки-ферменты)
елок-репрессор
Е1 Е2 Е3
(белки-ферменты)





 продукт
метаболит1
метаболит2 субстрат
продукт
метаболит1
метаболит2 субстрат


инактивация репрессора
У эукариот экспрессия генов не исследована. Причина – серьезные препятствия:
-организация генетического материала в форме хромосом.
- у многоклеточных организмов клетки специализированы и поэтому часть генов выключена.
- наличие гистоновых белков, в то время как у прокариот - «голая» ДНК.
№ 27. Геномные мутации - это изменение числа хромосом в геноме клетки.
Механизмы возникновения:
1. Нерасхождение хромосом во время мейоза или во время митотического деления соматических клеток на этапах дробления зиготы. В последнем случае наблюдается явление под названием мозаицизм: в организме развиваются клетки трех популяций — нормальные клетки, клетки-трисомики и клетки-моносомики.
2. Потеря отдельной хромосомы вследствие так называемого "хромосомного отставания" во время митотического деления клеток зародыша. При этом в организме появляется две популяции клеток: нормальные и клетки-моносомики.
3. Полиплоидизация — увеличение количества наборов хромосом (более двух). Отсутствие цитокинеза.
4. Изменение количества хромосом без изменения количества наследственного материала — робертсоновские перестройки: а) центрическое слияние, б) центрическое деление хромосом.
Полиплоидия - наследственное изменение, связанное с кратным увеличением основного числа хромосом в клетках организма. Полиплодия широко распространена у растений (более крупные размеры, повышенное содержание ряда веществ, лучшая устойчивость к неблагоприятным условиям внешней среды и т.п.) Различают два типа полиплоидов: аутополиплоиды - кратное увеличение наборов хромосом одного и того же вида и аллополиплоиды - соединение наборов хромосом разных организмов.
Анеуплоидия (гетероплоидия) - изменение генома, связанное с добавлением к набору одной или более хромосом или с их утратой. Трисомия — наличие трёх гомологичных хромосом в кариотипе. Моносомия — наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная совместимая с жизнью моносомия у человека — по хромосоме X — приводит к развитию синдрома Шерешевского—Тернера (45,Х0).
Значение. Возникновение геномных мутаций у млекопитающих известно только в качестве аномалий и приводит к гибели эмбриона на ранних стадиях развития. Однако изредка вследствие нерасхождения половых хромосом либо при одном, либо при двух делениях мейоза, возникает так называемая трисомия - XXY; а также моносомия — ХО, вызывающая серьезные гормональные нарушения у плода. Эволюция. В ряде случаев они позволяют увеличить набор генов, повышая генетическое богатство популяции.
Вопроса № 28 нет в билетах.
№ 29 Эволюция генома. Связана с нарастающим увеличением количества ДНК. Это увеличение наблюдается в процессе прогрессивной эволюции эукариот. На фоне такого увеличения большая часть ДНК является молчащей. Наиболее резко размер генома изменяется в результате полиплоидизации. По-видимому, в ходе эволюции в результате накопления мутаций и ивергенции нуклеотидных последовательностей полиплоидизация сопровождалась переходом к диплоидному состоянию. Если бы увеличение объема генома происходило только в результате полиплоидизации, то в природе должно было бы наблюдаться скачкообразное изменение его размеров. На самом деле этот процесс демонстрирует плавное увеличение содержания ДНК в геноме.
Значение имеют хромосомные перестройки: такие, как дупликации, делении и транслокации. Они обусловливают повторение, утрату некоторых последовательностей в составе хромосомы или перенос их в другие хромосомы.
Важный механизм - амплификация нуклеотидных последовательностей, которая заключается в образовании их копий, что приводит к возникновению повторяющихся участков ДНК. Особенностью генома эукариот является наличие таких повторов в большом количестве, свидетельствующее о существенном вкладе механизма амплификации в увеличение размеров наследственного материала. Причины амплификации: неравный кроссинговер → многократные дупликации отдельных участков ДНК. Возможна амплификация путем вырезания фрагмента с последующей его репликацией вне хромосомы и встраиванием копий в другие хромосомы. Пример: около 1100 млн. лет назад произошла дупликация гена - предшественника, давшая начало гемоглобиновым и миоглобиновым генам. Позднее, около 500 млн. лет назад, на ранней стадии эволюции позвоночных произошла дупликация, давшая начало двум (α и β) семействам глобиновых генов, сопровождавшаяся транслокацией. Примерно 200 млн. лет назад очередная дупликация привела к возникновению в семействе β-глобиновых генов генов β- глобинов плодов и взрослых. Около 100 млн. лет назад произошло образование ε- и γ-глобиновых генов и, наконец, 40 млн. лет назад появились δ- и β-глобиновые гены.
Подвижные генетические элементы - транспозоны. Они представляют собой автономные единицы, несущие в нуклеотидной последовательности информацию о структуре особых белков, которые обеспечивают их способность к перемещению из одного участка генома в другой. Такое перемещение - транспозиция - может происходить в строго определенные участки хромосом, узнаваемые этими специфическими белками. Транспозиция предполагает репликацию нуклеотидной последовательности подвижного генетического элемента и встраивание копии в ДНК-мишень с сохранением другой копии в прежнем месте. В результате таких перемещений могут активироваться ранее не активные гены, и наоборот.
Роль горизонтального переноса генетического материала в эволюции генома. Может происходить и между организмами разных видов, одновременно существующими на Земле. В настоящее время доказана возможность изменения наследственных свойств у бактерий путем введения в бактериальную клетку чужеродной ДНК при конъюгации или с помощью фагов. Чужеродную ДНК можно ввести и в эукариотическую клетку, где она будет сохраняться как внехромосомный элемент или интегрироваться в геном и экспрессироваться. Гены могут переходить от одного эукариотического организма к другому и даже от эукариот к прокариотам, хотя это происходит крайне редко. Ведущая роль в горизонтальном переносе генетической информации принадлежит вирусам. В настоящее время широко обсуждается роль ретровирусов, наследственный материал которых представлен молекулой РНК, в перенесении информации от клетки к клетке. Но подобное явление происходит крайне редко, так как необходимость взаимного приспособления генов в геноме ограничивает возможность встраивания в него чужеродных функциональных последовательностей.
Секвенирование генома. Секвенирование биополимеров (белков и нуклеиновых кислот — ДНК и РНК) — определение их первичной аминокислотной или нуклеотидной последовательности.
Чтение первого генома человека заняло 15 лет. Вслед за этим последовали секвенции–последовательности отдельных хромосом и создание машин-секвенаторов следующего поколения.
№30 Размножение – приспособление организмов к продолжению жизни. Размножение связано на молекулярном уровне с репликацией ДНК.
Бесполое размножение характеризуется тем, что новая особь развивается из неполовых, соматических клеток. Участвует только одна исходная особь. В этом случае организм может развиться из одной клетки, а возникшие потомки по своим наследственным признакам идентичны материнскому организму. Бесполое размножение широко распространено среди растений и значительно реже встречается у животных. Многие простейшие размножаются путем обычного митотического деления клетки - путем деления материнской клетки пополам (бактерии, эвглены, амебы, инфузории). Спорообразование. Оно заключается в том, что клетка распадается на большое число особей в результате многократного деления ее ядра (малярийный плазмодий). Многоклеточные организмы также способны к спорообразованию: у грибов, водорослей, мхов и папоротникообразных споры и зооспоры образуются в специальных органах — спорангиях и зооспорангиях. Один из способов – почкование.
Половое. Преемственность поколений и увеличение численности особей осуществляется с помощью специализированных половых клеток — гамет: женских — яйцеклеток и мужских — сперматозоидов. Созревшие половые клетки при слиянии образуют зиготу, из которой развивается новый дочерний организм. По достижении половой зрелости новый организм в свою очередь производит гаметы, которые дают начало следующим потомкам.
Преимущества полового размножения:
Способность популяции к более быстрому изменению.
Облегчение видообразования.
Большое генетическое разнообразие в потомстве облегчает адаптацию к непредсказуемым условиям среды.
Это обусловлено тем, что генотип потомков возникает путем комбинации генов, принадлежащих обоим родителям. При партеногенезе, когда развитие нового организма происходит только из яйцеклетки, в ней возникает новая комбинация генов в результате кроссинговера и независимого комбинирования хромосом.
В половых железах развиваются половые клетки — гаметы. Мужские гаметы созревают в мужских половых железах — семенниках; этот процесс называется сперматогенезом. Женские гаметы созревают в яичниках в процессе овогенеза. В процессе образования половых клеток — как сперматозоидов, так и яйцеклеток — выделяют ряд стадий: зону размножения, зону роста и зону созревания; в зоне созревания гаметы окончательно формируются путем мейоза.
№31. Гаметогенез. Сперматогенез. Морфологически семенник состоит из множества семенных канальцев. Дольчатое строение. Между семенными канальцами – клетки Лейдинга (начинают работу в 12-14 лет) синтезируют тестостерон – развитие вторичных половых признаков. Семенник очень рано становится эндокринным органом, под влиянием андрогенов происходит формирование мужских половых органов. Семенной каналец имеет зоны:
-размножения,
-роста,
-созревания и формирования.
Существуют одноименные периоды роста. Зона размножения в наружной части семенника. Клетки округлые, цитоплазмы много, ядро большое – сперматогонии. Они размножатся митозом, и семенник увеличивается в размерах до полового созревания, после – делятся только стволовые клетки. Запас клеток не уменьшается и семенник тоже не уменьшается. В зоне размножения 2n2c.следующая фаза – роста. Увеличивается размер ядра, цитоплазмы, идет репликация ДНК (интерфаза 1), клетки – сперматоциты первого порядка 2n4c. Эти клетки вступают в зону формирования и созревания у семенных канальцев. Мейоз состоит из 2 митотических делений, после первого деления n2c, после второго – nc.
Овогенез (яичники). Половые железы закладываются на 2м месяце эмбрионального развития. У человека очень рано закладывается желточный мешок (функция формирования первичных половых клеток, обеспечение питательными веществами). Половые клетки (первичные) мигрируют в развивающуюся половую железу, а желточный мешок дегенерирует. В эмбриогенезе яичники не активны. Формирование женских половых клеток пассивное. Первичные половые клетки – овогонии, они делятся. Формируются овоциты первого порядка. Период деления оканчивается к 7му месяцу эмбриогенеза – 7000000 первичных клеток. 400-500 созревают в течение жизни, остальные невостребованы. Развитие яйцеклеток у человека блокируется в профазе первого мейотического деления (на стадии диплотены). С наступлением половой зрелости овоцит увеличивается в размере, растет и размер желтка. Накапливаются пигменты, происходят биохимические и морфологические изменения. Каждый овоцит окружается мелкими фолликулярными клетками, созревающими в фолликуле. Яйцеклетка, созревая, приближается к периферии. Фолликулярная жидкость окружает её на всех этапах. Фолликул разрывается. Яйцеклетка попадает в брюшную полость. Затем в воронку яйцевода. Продолжение мейоза в 2/3 яйцевода в результате контакта яйцеклетки со сперматозоидом.
При мейозе идет распределение хромосом. В результате 4 ядра. Происходит конъюгация хромосом (за счет высоко повторяющихся последовательностей ДНК в 1ген). Каждое из 4х ядер при гаметогенезе получает только 1 хроматиду из пары. В результате мейоза при сперматогенезе из каждого спермацита первого порядка получаются 4 хроматиды и формируются 4 сперматозоида. Из одного овоцита первого порядка образуются 2 ядра с гаплоидным набором хромосом. Одно из них, с большим количеством цитоплазмы (т.к. при цитокинезе разделение идет неравномерно) и другое – редукционное (направляющее) тельце. При последующем делении образуются яйцеклетка и направляющее тельце. При овогенезе из каждого овоцита формируется 1 яйцеклетка и 3 направляющих тельца, которые дегенерируют и исчезают. В яйцеклетке есть все необходимые запасы питательных веществ.
Мейоз – способ деления диплоидных клеток с образованием из одной материнской диплоидной клетки четырех дочерних гаплоидных клеток. Мейоз состоит из двух последовательных делений ядра и короткой интерфазы между ними.
Первое деление состоит из профазы I, метафазы I, анафазы I и телофазы I.
В профазе I парные хромосомы, каждая из которых состоит из двух хроматид, подходят друг к другу (этот процесс называется конъюгацией гомологичных хромосом), перекрещиваются (кроссинговер), образуя мостики (хиазмы), затем обмениваются участками. При кроссинговере осуществляется перекомбинация генов. После кроссинговера хромосомы разъединяются.
В метафазе I парные хромосомы располагаются по экватору клетки; к каждой из хромосом прикрепляются нити веретена деления.
В анафазе I к полюсам клетки расходятся двухроматидные хромосомы; при этом число хромосом у каждого полюса становится вдвое меньше, чем в материнской клетке.
Затем наступает телофаза I – образуются две клетки с гаплоидным числом двухроматидных хромосом; поэтому первое деление мейоза называют редукционным.
После телофазы I следует короткая интерфаза (в некоторых случаях телофаза I и интерфаза отсутствуют). В интерфазе между двумя делениями мейоза удвоения хромосом не происходит, т.к. каждая хромосома уже состоит из двух хроматид.
Второе деление мейоза отличается от митоза только тем, что его проходят клетки с гаплоидным набором хромосом; во втором делении иногда отсутствует профаза II.
В метафазе II двухроматидные хромосомы располагаются по экватору; процесс идет сразу в двух дочерних клетках.
В анафазе II к полюсам отходят уже однохроматидные хромосомы.
В телофазе II в четырех дочерних клетках формируются ядра и перегородки (в растительных клетках) или перетяжки (в животных клетках). В результате второго деления мейоза образуются четыре клетки с гаплоидным набором хромосом (1n1c); второе деление называют эквационным (уравнительным) (рис. 18). Это – гаметы у животных и человека или споры у растений.
Значение мейоза состоит в том, что создается гаплоидный набор хромосом и условия для наследственной изменчивости за счет кроссинговера и вероятностного расхождения хромосом
У большинства диких животных сперматогенез происходит лишь в определенные периоды года. В промежутках между ними в канальцах семенников содержатся лишь сперматогонии. Но у человека животных сперматогенез происходит в течение всего года.
У многих животных овогенез и созревание яиц совершаются лишь в определенные сезоны года. У женщин обычно ежемесячно созревает одно яйцо, а за весь период половой зрелости около 400 яиц.
№32. Морфология половых клеток. Функции сперматозоида – оплодотворение (стимуляция дальнейшего развития яйцеклетки), обеспечение генетической информацией мужского организма. Все сперматозоиды имеют жгутики, подвижны, небольшого размера (50-90 мкм у человека). Состоят из головки, шейки, средней части и хвостика. Головка -5мкм, шейка – 5. Головка сперматозоида почти полностью занята ядром, цитоплазмы мало, она в жидкокристаллическом состоянии (защита от вредных явлений – ионизирующего излучения). Находится по периферии ядра. На конце головки – акросома с видоизмененным комплексом Гольджи; Ферменты: гиалуронидаза, муциназа. В плазматической мембране – проакрозин, который превращается в акрозин, проходя по половым путям самки (происходит отщепление ингибитора). Функция акрозина – отщепление фолликулярных клеток, отщепление блестящей оболочки. Шейка содержит пару центриолей. Микротрубочки одной из них удлиняются, образуется основная нить хвостика. В шейке много митохондрий, расположенных по спирали.
Органеллы движения – жгутики, способны к биению только при смешивании с секретом предстательной железы при семяизвержении. При нарушении функций предстательной железы – мужская стерильность.
Яйцеклетка. Функции: передает зародышу половину его будущего хромосомного набора; во время оплодотворения яйцеклетка приносит гораздо больше цитоплазмы; яйцеклетка снабжает зародыш пищевыми запасами до начала его собственного питания.
Размеры яйцеклеток намного больше размеров сперматозоидов(130-150 мкм у человека). В зрелой яйцеклетке запасаются все материалы, которые обеспечивают начальные стадии развития зародыша. Если сперматозоид, созревая, старается избавиться от цитоплазмы, яйцеклетка, наоборот, стремится увеличить ее количество. Есть рибосомы, р-РНК, т-РНК, морфогенетические факторы. Многие белки синтезируются в печени, жировом теле, а затем транспортируются в яйцеклетку. Яйцеклетка имеет плазматическую мембрану. Во время оплодотворения плазматическая мембрана контролирует поступление многих ионов (например, натрия). К ней прилегает желточная оболочка (гликопротеины – специфическое прикрепление сперматозоида своего вида к соответствующей яйцеклетке), часто прозрачна, яйцеклетка окружена слоем клеток лучистого яйца – фолликулярными питающими клетками. Для оплодотворения сперматозоид должен пройти сквозь все оболочки.
Наследственный материал, приносимый яйцеклеткой и сперматозоидом по размеру одинаков.
Процесс образования яйцеклеток в яичниках - овогенез, оогенез. Сперматозоиды образуются в семенниках, процесс носит название сперматогенеза. Те и другие клетки образуются по-разному, но есть некоторые общие черты.
№33. Оплодотворение. Зрелая яйцеклетка может быть оплодотворена на протяжении небольшого отрезка времени (12-24 часа после овуляции у человека, у рыб и амфибий - через несколько минут после откладки). После овуляции яйцеклетка претерпевает серьезные клеточные изменения. Для оплодотворения очень важен срок жизни. Совокупность изменений яйцеклетки после овуляции – старение яйцеклетки, через 24 часа – перезрелая яйцеклетка – оплодотворение невозможно. Если происходит оплодотворение в данном случае, то невозможна имплантация. Возможны аборты. Ежемесячно созревает только 1 яйцеклетка.
Жизнеспособность сперматозоидов зависит от многих факторов. В щелочной среде они активны, но быстро погибают в кислой среде. Путь от влагалища до 2/3 яйцевода составляет 2-3 часа. В верхней трети влагалища происходит подщелачивание среды (т. к. кислотность высокая) из-за семенной жидкости. В матку проникают только сперматозоиды. Они сохраняют большую подвижность при пониженной температуре.
В процессе оплодотворения выделяют 3 этапа.
1) активация яйцеклетки
2) проникновение сперматозоида в яйцеклетку
3) слияние ядер.
В ходе оплодотворения сперматозоид преодолевает все оболочки яйцеклетки. Как только происходит контакт сперматозоида с яйцеклеткой – завершается мейоз яйцеклетки, в т.ч. она выделяет фертилизин. Сперматозоид выделяет антифертилизин – происходит приклеивание. В это время акрозин разрушает оболочки, выделяются ферменты. У человека сперматозоид проникает по-разному: либо только головка, либо шейка и головка. После контакта сперматозоида и яйцеклетки у моноспермных в течение 1-3 минут происходит кортикальная реакция – образуется оболочка оплодотворения. Если несколько сперматозоидов – гибель клетки. Ядро сперматозоида набухает, разрыхляется, образуется мужской пронуклеус. Ядро яйцеклетки превращается в женский пронуклеус. Идет репликация ДНК. Пронуклеусы идут к центру, ядерные оболочки исчезают, происходит слияние пронуклеусов (кариогамия), образуется зигота (2n4c). Через сутки начинается дробление. Возможно развитие эмбриона без участия мужской гаметы (механическое, тепловое, химическое воздействие) – партеногенез.
Партеногенез - форма полового размножения, развитие яйцеклетки без оплодотворения. Свойствен многим беспозвоночным животным (дафнии, коловратки, тли, пчелы и др.) и многим семенным и споровым растениям.
Существует два типа партеногенеза: облигатный (т.е. обязательный) и факультативный. Первый свойствен видам, у которых самцов либо нет совсем, либо они редки и не способны функционировать. К таким видам относятся некоторые тли, палочники, сверчки, бабочки; популяции без самцов изредка встречаются у рыб, например у серебряного карася. При факультативном партеногенезе яйца могут развиваться как партеногенетически, так и в результате оплодотворения, причем партеногенетическое размножение может преобладать в условиях, когда слишком редки контакты разнополых особей, например на границе ареала распространения вида.
Особые формы партеногенеза - гиногенез и андрогенез. Гиногенез – развитие из яйцеклетки. При гиногенезе сперматозоид проникает и погибает (только женские организмы, т.к. только женский геном). Андрогенез – развитие из сперматозоида. При нем ядро яйцеклетки погибает (только мужской геном). Очень быстро такой геном перерождается в злокачественную опухоль - хорионэпителиому.
Типы определения пола.
- прогамный. Пол будущего организма определяется в ходе гаметогенеза у родительских особей.
- сингамный. Пол будущего организма определяется в момент слияния половых клеток.
- эпигамный. Пол будущего организма определяется в процессе онтогенеза. У человека имеет место переопределение пола (при патологии) – хотя истинного нет.
№ 34. Предмет, задачи, методы генетики. Генетика — наука о законах наследственности и изменчивости организмов и методах управления ими.
Предмет генетики - изучение структуры и механизма функционирования генетического материала в живых системах и их изменчивости.
Важнейшая задача Г. — разработка методов управления наследственностью и наследственной изменчивостью для получения нужных человеку форм организмов или в целях управления их индивидуальным развитием. Задачи генетики вытекают из установленных общих закономерностей наследственности и изменчивости. К этим задачам относятся исследования: 1) механизмов хранения и передачи генетической информации от родительских форм к дочерним; 2) механизма реализации этой информации в виде признаков и свойств организмов в процессе их индивидуального развития под контролем генов и влиянием условий внешней среды; 3) типов, причин и механизмов изменчивости всех живых существ; 4) взаимосвязи процессов наследственности, изменчивости и отбора как движущих факторов эволюции органического мира.
Методы:
1. Гибридологический анализ, Заключающийся в использовании системы скрещиваний для установления характера наследования признаков и генетических различий изучаемых организмов. Гибридологический анализ, дополненный после работ Г. Менделя рядом специфических методов и приемов исследования наследственности, вошел в качестве важнейшей составной части в генетический анализ — основной метод генетики.
2. Цитологический метод — изучение структур клеток в связи с размножением организмов и передачей наследственной информации. На основе этого метода при использовании новейших способов изучения хромосомных структур возникла новая наука — цитогенетика.
3. Онтогенетический метод Используется для изучения действия генов и проявления их в индивидуальном развитии организмов — онтогенезе в разных условиях внешней среды.
4. Статистический метод, С помощью которого изучают статистические закономерности наследственности и изменчивости организмов.
История генетики
1865 Грегор Мендель делает доклад Опыты над растительными гибридами.
1903 Высказано предположение о том, что хромосомы являются носителями наследственности.
1905 Уильям Бэтсон вводит термин генетика.
1908 Открыт закон Харди — Вайнберга.
1910 Томас Хант Морган доказывает, что гены расположены в хромосомах.
1956 Jo Hin Tjio и Алберт Леван впервые верно устанавливают Хромосомное число человека: 46 хромосом в диплоидном наборе.
1961 Выяснено, что генетический код состоит из триплетов.
1977 ДНК секвенирована впервые.
1989 Впервые секвенирован ген человека.
2003 (14 апреля) Проект «Геном человека» успешно завершён: 99 % генома секвенировано с точностью 99,99 %.
Н. И. Вавилов. Создал учение о мировых центрах происхождения культурных растений. Открыл закон гомологических рядов. Внёс существенный вклад в разработку учения о биологическом виде.
Н. К. Кольцов. Один из основателей генетики в России. Создатель Института экспериментальной биологии в Москве (лето 1917 года). Он впервые ясно сформулировал матричный принцип репродукции молекулярной структуры наследственного материала (хромосомы как наследственные молекулы).
А. С. Серебровский. Показал сложную структуру гена, ввел в науку термин «генофонд». Предложил принципиально новый метод борьбы с насекомыми-вредителями, основанный на массовом выпуске самцов вредных видов с генетическими аномалиями (1940).
С. С. Четвериков. Объединил в единой концепции идеи менделизма и дарвиновской теории эволюции. Доказал, что мутации (геновариации) в природных популяциях животных не исчезают, могут накапливаться в скрытом (гетерозиготном) состоянии и давать материал для изменчивости и естественного отбора.
№ 35. Генотип - совокупность всех генов организма (генетическая конституция). Генотип постоянно испытывает воздействие внешней среды, он взаимодействует со средой, что приводит к формированию всех признаков и свойств организма.
Фенотип – все признаки организма, формирующиеся в результате взаимодействия генотипа и среды. (Иогансен – 1803год) свойства любого организма зависят от генотипа и от среды, поэтому формирование организма – результат взаимодействия генетических факторов и факторов внешней среды.
Признак – единица морфологической, физиологической, биохимической, иммунологической, клинической и любой другой дискретности организмов (клеток), т.е. отдельное качество или свойство, по которому они отличаются друг от друга.
Аллельные гены - различные формы одного и того же гена, расположенные в одинаковых участках (локусах) гомологических хромосом. Аллели определяют варианты развития одного и того же признака. В нормальной диплоидной клетке могут присутствовать не более двух аллелей одного локуса одновременно. В одной гамете два аллеля находиться не могут. Форма взаимодействия между аллельными генами может быть не только в виде доминирования (полное подавление проявления одного гена другим).
Взаимодействия неаллельных генов.
1. Комплементарность. Этот вид взаимодействия генов заключается в том, что при наличии двух доминантных аллелей разных генов появляется новый признак, то есть для появления нового признака у организма должен быть генотип АВ.
2. Эпистаз. При эпистатическом взаимодействии одна пара генов может подавлять действие другой пары генов.
3. Полимерия. Многие признаки определяются несколькими парами генов. Это характерно, в основном, для количественных признаков, таких как яйценоскость у кур, жирность молока у коров.
4. Плейотропное действие гена. При плейотропном действии гена один ген определяет развитие или влияет на проявление нескольких признаков.
ГОМОЗИГОТНОСТЬ — состояние организма, при к-ром гомологичные хромосомы несут одну и ту же аллель того или иного гена. Самооплодотворяющиеся организмы практитически гомозиготны.
Гетерозиготность - состояние, при котором гомологичные хромосомы несут разные формы (аллели) того или иного гена.
Гемизиготность —состояние, связанное с тем, что у организма один или несколько генов не парные, т. е. не имеют аллельных партнеров.
№ 36. Закономерности наследования при моногибридном скрещивании.
Моногибридное скрещивание — скрещивание форм, отличающихся друг от друга по одной паре альтернативных признаков.
Первый закон Менделя (единообразия гибридов) – при скрещивании гомозиготных родительских особей, которые отличаются по одной паре аллельных признаков, все гибриды первого поколения единообразны по фенотипу и генотипу.
Второй закон Менделя (расщепления гибридов второго поколения) – при моногибридном скрещивании гетерозиготных организмов у гибридов второго поколения происходит расщепление по фенотипу в отношении 3:1 и по генотипу – 1:2:1

№ 37. Дигибридное и полигибридное скрещивание. Дигибридное скрещивание - скрещивание организмов, различающихся по двум парам альтернативных признаков.
Третий закон Менделя (независимого наследования признаков) – при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.
Закон проявляется, как правило, для тех пар признаков, гены которых находятся в не гомологичных хромосомах. Если обозначить буквой и число аллельных пар в негомологичных хромосомах, то число фенотипических классов будет определяться формулой 2n, а число генотипических классов - 3n. При неполном доминировании количество фенотипических и генотипических классов совпадает.

Цитологические основы законов Менделя базируются на:
1) парности хромосом (парности генов, обусловливающих возможность развития какого-либо признака)
2) особенностях мейоза (процессах, происходящих в мейозе, которые обеспечивают независимое расхождение хромосом с находящимися на них генами к разным полюсам клетки, а затем и в разные гаметы)
3) особенностях процесса оплодотворения (случайного комбинирования хромосом, несущих по одному гену из каждой аллельной пары)
Формула расщепления по обоим признакам. 9:3:3:1
Вопрос №37.
Дигибридное и полигибридное скрещивание. Закон независимого комбинирования генов и его цитологические основы. Общая формула расщепления при независимом наследовании.
Дигибридное скрещивание - скрещивание двух особей, отличающихся друг от друга по двум парам альтернативных признаков.
Полигибридное скрещивание - скрещивание двух особей, отличающихся друг от друга по двум и более парам альтернативных признаков.
Закон независимого комбинирования генов. Третий закон Менделя (независимого наследования признаков) – при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум и более парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях.
Закон проявляется, как правило, для тех пар признаков, гены которых находятся в негомологичных хромосомах.
Цитологические основы: случайное комбинирование хромосом при оплодотворении, несущих по одному гену из каждой аллельной пары.
Общая формула расщепления при независимом наследовании. По фенотипу: 9 (А-В-):3(А-вв):3(ааВ-):1(аавв).
Вопрос №38.
Множественные аллели. Наследование групп крови человека системы AB0.
Множественные аллели – различные состояния (три или более) одного и того же локуса хромосомы, возникшие в результате мутаций. Множественный аллелизм - наличие у гена множественных аллелей. Примером множественного аллелизма может служит система групп крови АВО.
Наследование групп крови. В локусе гена АВО возможны три варианта (аллеля) - 0, A и B, которые экспрессируются по аутосомно-кодоминантному типу. Это означает, что у лиц, унаследовавших гены А и В, экспрессируются продукты обоих этих генов, что приводит к образованию фенотипа АВ (IV). Фенотип А (II) может быть у человека, унаследовавшего от родителей или два гена А, или гены А и 0. Соответственно фенотип В (III) - при наследовании или двух генов В, или В и 0. Фенотип 0 (I) проявляется при наследовании двух генов 0.
Вопрос №39.
Взаимодействие неаллельных генов: комплементарность, эпистаз, полимерия, модифицирующее действие.
Большинство признаков и свойств организма являются результатом действия не одной пары аллельных генов, а нескольких неаллельных генов или их продуктов. Поэтому эти признаки называют сложными. Сложным признаком может быть четвертичная структура белка, образуемая разными пептидными цепями, которые детерминируются разными (неаллельными) генами.
Полимерия. Если гены располагаются, каждый в своем отдельном локусе, но их взаимодействие проявляется в одном и том же направлении – это полигены. Один ген проявляет признак незначительно. Полигены дополняют друг друга и оказывают мощное действие – возникает полигенная система – т.е. система является результатом действия одинаково направленных генов. Совместное действие полигенов обусловливает различную экспрессивность — степень выраженности признака, зависящую от дозы соответствующих аллелей. По полимерному типу взаимодействия у человека определяется интенсивность окраски кожных покровов, зависящая от уровня отложения в клетках пигмента меланина.
Комплементарность – явление, при котором 2 неаллельных гена, находясь в генотипе, одновременно приводят к формированию нового признака. Если присутствует один из пары – проявляется он. Примером служат группы крови у человека. Комплементарность может быть доминантная и рецессивная.
Эпистаз – такое взаимодействие генов, когда ген одной аллельной пары маскируется действием другой аллельной пары. Это обусловлено тем, что ферменты катализируют разные процессы клетки, когда на одном метаболическом пути действуют несколько генов. Действие их должно быть согласовано во времени.
Модифицирующее (гены-модификаторы) — усиление или ослабление действия главных генов действием других, неаллельных им генов. Ген-модификатор (1919, Бриджес). Теоретически любой ген может взаимодействовать с другими генами, а значит, и проявлять модифицирующее действие, но некоторые гены – модификаторы в большей степени. Они часто не имеют собственного признака, но способны усиливать или ослаблять проявление признака, контролируемого другим геном. В формировании признака кроме основных генов проявляют свое действие и модифицирующие гены.
Примеры:
окраска млекопитающих – белая, черная + модификаторы;
Фенилкетонурия. У больных есть мутация, которая выключает фермент – фенилаланин – гидролазу. Поэтому фенилаланин не превращается в тирозин. В результате в крови количество фенилаланина повышается.
Вопрос №40.
Хромосомная теория наследственности. Сцепление генов. Группы сцепления. Кроссинговер как механизм, определяющий нарушения сцепленного гена.
Хромосомная теория наследственности.
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
3. Гены расположены в хромосомах в определенной линейной последовательности;
4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом;
6. Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость); 7. Каждый вид имеет характерный только для него набор хромосом – кариотип.
Кроссинговер нарушает сцепленность этих генов.
Полное и неполное сцепление генов. Гены в хромосомах имеют разную силу сцепления. Сцепление генов может быть: полным, если между генами, относящимися к одной группе сцепления, рекомбинация невозможна и неполным, если между генами, относящимися к одной группе сцепления, возможна рекомбинация.
Вопрос №41.
Наследование. Типы наследования. Особенности аутосомного, X-сцепленного и голандрического типов наследования. Полигенное наследование.
Наследование – передача информации от одного поколения к другому.
Типы наследования: - аутосомное: доминантное, рецессивное, промежуточное.
- сцепленное с полом: X-сцепленное и голандрическое (Y-сцепленное).
Аутосомно-доминантный тип наследования. Признак проявляется и в гомозиготном, и в гетерозиготном состоянии. Это способность свертывать язык в трубочку и «свисающая» (свободная) мочка уха. Альтернативой последнему признаку является срощенная мочка – признак рецессивный. Еще одна наследственная аномалия у человека, обусловленная аутосомно-доминантным геном, – многопалость, или полидактилия.
Аутосомно-рецессивный тип наследования. Например, голубой цвет глаз проявляется у людей, гомозиготных по соответствующему аллелю. Признак рыжих волос, определяющий еще и характер пигментации кожи, также является рецессивным по отношению к не рыжим волосам и проявляется только в гомозиготном состоянии.
Наследование сцепленное с полом.
Локализацию генов в половых хромосомах называют сцеплением генов с полом. Наследование определяемых ими признаков называют наследованием, сцепленным с полом. Например, Х-хромосома человека содержит доминантный ген Н,
пределяющий свертывание крови. У человека, рецессивно гомозиготного по этому признаку, развивается тяжелое заболевание гемофилия. Так как в клетках женщин две Х-хромосомы, то наличие в одной из них гена h не влечет за собой заболевания, так как во второй из них присутствует доминантный ген Н. В клетках мужчин есть только одна Х-хромосома. Если в ней присутствует ген h, то у мужчины разовьется гемофилия, так как Y-хромосома не гомологична Х-хромосоме и в ней нет гена Н или h.
Голандрическое наследование – гипертрихоз ушной раковины («волосатые уши»).
Вопрос №42.
Количественная и качественная специфика проявления генов в признаках: пенетрантность, экспрессивность, плейотропность, генокопии.
Пенетрантность – количество особей (%), проявляющих в фенотипе данный ген, по отношению к количеству особей, у которых этот признак мог бы проявиться.
Экспрессивность – это степень проявления фенотипа. Количественно ее степень определяют, используя статистический показатель.
Плейотропность - это множественное действие генов. Это когда один ген отвечает за много признаков.
Вопрос №43.
Изменчивость. Формы изменчивости: модификационная и генотипическая, их значение в онтогенезе и эволюции.
Изменчивость – свойство живых организмов существовать в разных формах. Групповая и индивидуальная изменчивость – классификация по эволюционному значению. Изменчивость, реализованная группой организмов, называется групповой, у одного организма или группы его клеток – индивидуальная.
По характеру изменения признаков и механизму:
--фенотипическая
- случайная
- модификационная
--генотипическая
- соматическая
- генеративная (мутационная, комбинативная)
а) генная
б) хромосомная
в) геномная
Модификационная изменчивость отражает изменение фенотипа под воздействием факторов внешней среды.
Соматическая изменчивость не передается по наследству.
Комбинативная изменчивость – результат независимого расхождения хромосом в процессе мейоза, оплодотворения, кроссинговера с рекомбинацией генов. При комбинативной изменчивости происходит перекомбинация генов, возникает новый индивидуальный набор хромосом, а значит, новый генотип и фенотип.
Мутационная изменчивость - вид изменчивости, в результате которого могут появиться новые гены, которые могли раньше не встречаться. Происходит изменение генотипа и как следствие, изменяется фенотип.
Вопрос №44.
Фенотипическая изменчивость и её виды.
Фенотипическая изменчивость:
- случайная
- модификационная.
Модификационная изменчивость отражает изменение фенотипа под воздействием факторов внешней среды.
Модификации - фенотипические изменения, возникающие на основе одного и того же генотипа в разных условиях его реализации. Примером модификаций могут служить изменения содержания жира в молоке животных или массы тела в зависимости от их питания, изменения количества эритроцитов в крови в зависимости от парциального давления кислорода в воздухе, изменения темпа роста растений при разной освещенности и содержании минеральных веществ в почве.
Норма реакции - пределы модификационной изменчивости признака, допустимой при данном генотипе.
Частный случай фенотипической изменчивости – фенокопии. Фенокопии – вызванные условиями внешней среды фенотипические модификации, имитирующие генетические признаки. Под влиянием внешних условий на генетически нормальный организм копируются признаки совсем другого генотипа. У человека возникает заболевание витилиго (1% людей) – нарушение пигментации кожи. Генетический дефект есть у 30% болеющих, у остальных – профессиональное витилиго (воздействие на организм особых химических и отравляющих веществ).
Адаптивный характер модификационной изменчивости — приспособительная реакция организмов на изменения условий среды.
Вопрос №45
Фенотип. Фенотип как результат реализации наследственной информации (генотипа) в определённых условиях среды. Значение средовых и генотипических факторов в формировании патологически изменённого фенотипа человека.
Фенотип – все признаки организма, формирующиеся в результате взаимодействия генотипа и среды. (Иогансен – 1803год). Свойства любого организма зависят от генотипа и от среды, поэтому формирование организма – результат взаимодействия генетических факторов и факторов внешней среды. Фенотипическое проявление генотипа в зависимости от среды изменяется в пределах нормы реакции. От родителей потомки получают специфические типы химических реакций на разные условия среды. Совокупность всех химических реакций определят метаболизм – обмен веществ. У каждого человека свои особенности обмена веществ, которые передается от поколения к поколению, и подчиняются законам Менделя. Различия в обмене веществ реализуются в конкретных условиях среды на уровне синтеза белка.
Сами признаки не наследуются. Признаки развиваются на основе взаимодействия генотипа и среды. Наследуется только генотип, т.е. комплекс генов, который определяет норму биологической реакции организма, изменяющую проявление и выраженность признаков в разных условиях среды. Таким образом, организм реагирует на свойства внешней среды. Иногда один и тот же ген в зависимости от генотипа и от условий внешней среды по-разному проявляет признак или меняет полноту выраженности.
Вопрос №46
Комбинативная изменчивость, её механизмы. Значение комбинативной изменчивости в обеспечении генотипического разнообразия людей.
Комбинативная изменчивость – результат независимого расхождения хромосом в процессе мейоза, оплодотворения, кроссинговера с рекомбинацией генов. При комбинативной изменчивости происходит перекомбинация генов, возникает новый индивидуальный набор хромосом, а значит, новый генотип и фенотип. Для комбинативной изменчивости в системе людей большое значение имеет система браков. Самая простая – случайный подбор пар (панмиксия). Строго панмиксных популяций не существует, т.к. существуют ограничения: социальные, религиозные, индивидуальные, экономические и другие. Поэтому в популяциях людей имеют место отклонения от панмиксии в двух направлениях:
Люди, состоящие между собой в родстве, вступают в брак чаще, чем при случайном подборе – инбридинг – инбредные (кровнородственные браки).
Люди вступают в брак чаще при случайном подборе пар, чем при родственном бракосочетании – аутобридинг.
Вопрос №47
Генные болезни человека, механизмы их возникновения и проявления. Примеры.
Генные (или менделевские) болезни - это моногенно обусловленные патологические состояния, наследуемые в соответствии с законами Менделя. По локализации гена в хромосомах выделяют:
Аутосомно-рецессивные (фенилкетонурия, муковисцидозы, болезнь Эллерса — Данлоса).
Аутосомно-доминантные (болезнь Марфана, ахондроплазия).
Х-сцепленные (ломкость Х хромосомы - снижение умственных способностей
У-сцепленные
В зависимости от функциональной значимости первичных продуктов соответствующих генов генные болезни подразделяют на наследственные нарушения ферментных систем (энзимопатии), дефекты белков крови (гемоглобинопатии), дефекты структурных белков (коллагеновые болезни) и генные болезни с невыясненным первичным биохимическим дефектом. Энзимопатии. В основе энзимопатии лежат либо изменения активности фермента, либо снижение интенсивности его синтеза (галактоземия - нарушение метаболизма молочного сахара, мукополисахаридозы - нарушение расщепления полисахаридов, фенилкетонурия - нарушение обмена фенилаланина ).
Гемоглобинопатии. Это группа наследственных заболеваний, вызываемых первичным дефектом пептидных цепей гемоглобина и связанным с этим нарушением его свойств и функций. К ним относят метгемоглобинемии, эритроцитозы, серповидно-клеточную анемию, талассемии.
Коллагеновые болезни. В основе возникновения этих заболеваний лежат генетические дефекты биосинтеза и распада коллагена — важнейшего структурного компонента соединительной ткани. К этой группе относят болезнь Эллерса — Данлоса, наследующуюся как по аутосомно-доминантному, так и по аутосомно-рецессивному типу, болезнь Марфана, наследующуюся по аутосомно-доминантному типу.
Наследственные болезни с невыясненным первичным биохимическим дефектом. К этой группе принадлежит подавляющее большинство моногенных наследственных болезней. Наиболее распространенными являются следующие.
1. Муковисцидозы — наследуются по аутосомно-рецессивному типу. В основе патогенеза заболевания —наследственное поражение экзокринных желез и железистых клеток организма, выделение ими густого, измененного по составу секрета и связанные с этим последствия.
2. Ахондроплазия —наследуется по аутосомно-доминантному типу. Это заболевание костной системы, при котором наблюдаются аномалии развития хрящевой ткани преимущественно в эпифизах трубчатых костей и костях основания черепа.
3. Мышечные дистрофии (миопатии) —заболевания, связанные с поражением поперечно-полосатых и гладких мышц.
Вопрос №48
Хромосомные болезни человека, механизмы их возникновения и проявления. Примеры.
Эта группа заболеваний обусловлена хромосомными мутациями (изменяется структура отдельных хромосом, но их число не меняется). Структурные перестройки хромосом также, как правило, сопровождаются дисбалансом генетического материала (делеции, дупликации). Степень снижения жизнеспособности при хромосомных аберрациях зависит от количества недостающего или избыточного наследственного материала и от вида измененной хромосомы. Хромосомные изменения, приводящие к порокам развития, чаще всего привносятся в зиготу с гаметой одного из родителей при оплодотворении. При этом все клетки нового организма будут содержать аномальный хромосомный набор. Если хромосомные нарушения возникают в одном из бластомеров во время первых делений зиготы, образующейся из нормальных гамет, то развивается мозаичный организм, большая или меньшая часть клеток которого несет нормальный хромосомный набор.
Транслокационный синдром Дауна, при котором число хромосом в кариотипе формально не изменено и равно 46, так как дополнительная 21-я хромосома транслоцирована на одну из акроцентрических хромосом. При делении короткого плеча 5-й хромосомы развивается синдром кошачьего крика, при котором наблюдаются общее отставание в развитии, низкая масса при рождении, лунообразное лицо с широко расставленными глазами и характерный плач ребенка, напоминающий кошачье мяукание, причиной которого является недоразвитие гортани.
У носителей некоторых перицентрических инверсий нередко наблюдаются аномалии в виде умственной отсталости той или иной степени и пороков развития. Довольно часто такие перестройки наблюдаются в 9-й хромосоме человека, однако они существенно не влияют на развитие организма.
Вопрос №49.
Геномные болезни человека, механизмы их возникновения и проявления. Примеры.
Геномные болезни вызываются геномными мутациями. У человека описаны геномные мутации по типу полиплоидии, которые редко наблюдаются у живорожденных, а в основном обнаруживаются у абортированных эмбрионов и плодов и у мертворожденных. Основную часть хромосомных болезней составляют анэуплоидии, причем моносомии по аутосомам у живорожденных встречаются крайне редко.
Примеры: 47,хх+21, 47,ху+21 Синдром Дауна.Большой язык, не помещающийся в полости рта, специфический разрез глаз, умственная отсталость и т.д. Не способны к трудовой деятельности, требуют ухода и дорогостоящего лечения.Чем старше мать, тем выше вероятность рождения ребенка с этой патологией.
47,хху – синдром Клайнфельтера. Мужчина. Высокий рост, более длинные ноги, евнуховидное телосложение, недоразвитие половых органов, гинекомастия, у половины умственная отсталость.
45,х0 синдром Шерешевского-Тернера. Женщины. Фенотипические проявления – маленький рост, для многих характерна шейная складка, укорочены 4 и 5 пальцы, антимонголоидные глаза, абстрактное мышление отсутствует.
47,хх+13,47,ху+13 Синдром Патау. Характерны микроцефалия (недоразвитие головного мозга), резкая умственная отсталость, незарощение неба и губы. Полидактилия, повышенная гибкость суставов, недоразвитие глазного яблока, неправильно сформированные, низко посаженные ушные раковины, пороки внутренних органов. Такие дети не живут долго.
47,хх+18, 47,ху+18 Синдром Эдвардса.
Частота встречаемости у девочек в 3 раза выше, чем у мальчиков. Характерны множественные аномалии, грубые пороки, характерна грубая задержка роста, своеобразный свод черепа, пяткообразно нависающий затылок, короткая шея, расстояние между висками маленькое, ушная раковина деформирована, у половины на затылке избыточная кожа. Продолжительность жизни таких детей снижена.
Вопрос №50.
Болезни человека с наследственной предрасположенностью, механизмы их возникновения и проявления. Примеры.
Эта группа болезней отличается от генных болезней тем, что для своего проявления нуждается в действии факторов внешней среды. Среди них также различают моногенные, при которых наследственная предрасположенность обусловлена одним патологически измененным геном, и полигенные. Последние определяются многими генами, которые в нормальном состоянии, но при определенном взаимодействии между собой и с факторами среды создают предрасположение к появлению заболевания. Они называются мультифакториальными заболеваниями. Моногенные заболевания - это наследственно обусловленные патологические реакции на действие различных внешних факторов (лекарственных препаратов, пищевых добавок, физических и биологических агентов), в основе которых лежит наследственная недостаточность некоторых ферментов.
К таким реакциям могут быть отнесены наследственно обусловленная непереносимость сульфаниламидных препаратов. У генетически чувствительных индивидов нежелательные реакции могут вызывать некоторые компоненты пищи и пищевые добавки. Известна непереносимость у ряда людей молочного сахара — лактозы. Иногда отмечается наследственная устойчивость к действию биологических агентов. Например гетерозиготы HbA HbS устойчивы к заражению возбудителем тропической малярии.
К болезням с наследственной предрасположенностью, обусловленной многими генетическими и средовыми факторами, относятся такие заболевания, как псориаз, сахарный диабет, шизофрения. Этим заболеваниям присущ семейный характер.
Нередко предрасположенность к ряду заболеваний наблюдается у людей с определенным сочетанием различных генов. Так, у людей со II (А) группой крови чаще наблюдается рак желудка и кишечника, а также сахарный диабет, ишемическая болезнь сердца, ревматизм. У людей с I (0) группой крови чаще встречается язвенная болезнь желудка и двенадцатиперстной кишки.
Вопрос №51
Значение генетики для медицины. Человек как специфический объект генетического анализа. Методы изучения наследственности человека: близнецовый, генеалогический, цитогенетический, биохимический, популяционно-статистический, генетика соматических клеток, методы изучения ДНК.
Генетика человека – наука о наследственно обусловленных различиях людей и о нарушениях генетического материала. Генетика человека – основа медицинской генетики. Гибридологический метод генетического анализа к человеку не применим. Во-первых, у человека не может быть произведено искусственного направленного скрещивания в интересах исследователя. Во-вторых, низкая плодовитость делает невозможным применение статистического подхода при оценке немногочисленного потомства одной пары родителей. В-третьих, редкая смена поколений, происходящая в среднем через 25 лет, при значительной продолжительности жизни дает возможность одному исследователю наблюдать не более 3—4 последовательных поколений. Наконец, изучение генетики человека затрудняется наличием в его геноме большого числа групп сцепления генов (23 у женщин и 24 у мужчин), а также высокой степенью фенотипического полиморфизма. Зато у человека лучше, чем у других видов изучены биохимические, иммунологические, физиологические и другие реакции, а эти признаки детерминированы генами.