

Erik_Kandel_-_V_poiskakh_pamyati_angl
.pdf4-5 Cajal's four principles of neural organization.
ron. With rare exceptions, all nerve cells in the brain have a cell body that contains a nucleus, a single axon, and many fine dendrites.
The axon of a typical neuron emerges at one end of the cell body and can extend up to several feet. The axon often splits into one or more branches along its length; at the end of each of these branches are many tiny axon terminals. The several dendrites usually emerge on the opposite side of the cell body (figure 4-5-A). They branch extensively, forming a treelike structure that grows out from the cell body
and spreads over a large area. Some neurons in the human brain have as many as forty dendritic branches.
In the 1890s Cajal pulled his observations together and formulated the four principles that make up the neuron doctrine, the theory of neural organization that has governed our understanding of the brain ever since.
The first principle is that the neuron is the fundamental structural and functional element of the brain—that is, both the basic building block and the elementary signaling unit of the brain. Moreover, Cajal inferred that the axons and dendrites play quite different roles in this signaling process. A neuron uses its dendrites to receive signals from other nerve cells and its axon to send information to other cells.
Second, he inferred that the terminals of one neuron's axon communicate with the dendrites of another neuron only at specialized sites, later named synapses by Sherrington (from the Greek synaptein, meaning to bind together). Cajal further inferred that the synapse between two neurons is characterized by a small gap, now

called the synaptic cleft, where the axon terminals of one nerve cell—which Cajal called the presynaptic terminals—reach out to, but do not quite touch, the dendrites of another nerve cell (figure 4-5-B). Thus, like lips whispering very close to an ear, synaptic communication between neurons has three basic components: the presynaptic terminal of the axon, which sends signals (corresponding to the lips in our analogy); the synaptic cleft (the space between lips and ear); and the postsynaptic site on the dendrite that receives signals (the ear).
Third, Cajal inferred the principle of connection specificity, which holds that neurons do not form connections indiscriminately. Rather, each nerve cell forms synapses and communicates with certain nerve cells and not with others (figure 4-5-C). He used the principle of connection specificity to show that nerve cells are linked in specific pathways he called neural circuits; signals travel along these circuits in a predictable pattern.
Typically, a single neuron makes contact through its many presynaptic terminals with the dendrites of many target cells. In this way, a single neuron can disseminate the information it receives widely to different
target neurons, sometimes located in different regions of the brain. Conversely, the dendrites of a target nerve cell can receive information from the presynaptic terminals of a number of different neurons. In this way a neuron can integrate information from a number of different neurons, even those located in different areas of the brain.
Based on his analysis of signaling, Cajal conceived of the brain as an organ constructed of specific, predictable circuits, unlike the prevailing view, which saw the brain as a diffuse nerve net in which every imaginable type of interaction occurred everywhere.
With an amazing leap of intuition, Cajal arrived at the fourth principle, dynamic polarization. This principle holds that signals in a neural circuit travel in only one direction (figure 4-5-D). Information flows, from the dendrites of a given cell to the cell body along the axon to the presynaptic terminals and then across the synaptic cleft to the dendrites of the next cell, and so on. The principle of the one-way flow of signals was enormously important because it related all components of the nerve cell to a single function—signaling.
The principles of connection specificity and the one-way flow of signals gave rise to a logical set of rules that has been used ever since to map the flow of signals between nerve cells. Efforts to delineate neural circuits received a further boost when Cajal showed that such circuits in the brain and spinal cord contain three major classes of neurons, each with a specialized function. Sensory neurons, which are located in the skin and in various sense organs, respond to a specific type of stimulus from the outside world—mechanical pressure (touch), light (vision), sound waves (hearing), or specific chemicals (smell and taste)—and send this information to the brain. Motor neurons send their axons out of the brain stem and spinal cord to effector cells, such as muscle and gland cells, and control the activity of those cells. Interneurons, the most numerous class of neurons in the brain, serve as relays between sensory and motor neurons. Thus Cajal was able to trace the flow of information from sensory neurons in the skin to the spinal cord and from there to interneurons and to motor neurons that signal muscle cells to move (figure 4-6). Cajal derived these insights from work on rats, monkeys, and people.
In time, it became clear that each cell type is biochemically distinct
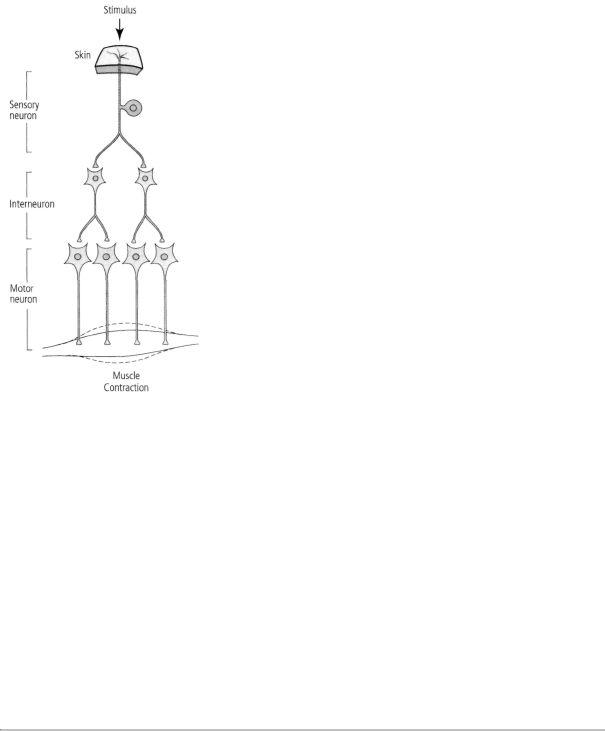
4-6 Three major classes of neurons, as identified by Cajal. Each class of neurons in the brain and spinal cord has a specialized function. Sensory neurons respond to stimuli from the outside world. Motor neurons control the activity of muscle or gland cells. Interneurons serve as relays between sensory and motor neurons.
and can be affected by distinct disease states. Thus, for example, sensory neurons from the skin and joints are compromised by a late stage of syphilis; Parkinson's disease attacks a certain class of interneurons; and motor neurons are selectively destroyed by amyotrophic lateral sclerosis and poliomyelitis. Indeed, some diseases are so selective that they affect only specific parts of the neuron: multiple sclerosis affects certain classes of axons; Gaucher's disease affects the cell body; fragile X syndrome affects dendrites; botulism toxin affects synapses.
For his revolutionary insights, Cajal received the Nobel Prize in Physiology or Medicine in 1906, together with Golgi, whose silver stain made Cajal's discoveries possible.
It is one of the strange twists of the history of science that Golgi, whose technical developments paved the way for Cajal's brilliant discoveries, continued to disagree vehemently with Cajal's interpretations and never subscribed to any aspect of the neuron doctrine.
Indeed, Golgi used the occasion of his Nobel Prize lecture to renew his attack on the neuron doctrine. He began by asserting once again that he had always been opposed to the neuron doctrine and that "this doctrine is generally recognized as going out of favor." He went on to say, "In my opinion, we cannot draw any conclusion, one way or the other, from all that has been said ... in being for or against the neuron doctrine." He argued further that the principle of dynamic polarization was wrong and that it was incorrect to think that the elements of a neural circuit connected in precise ways or that different neural circuits had different behavioral functions.

Until his death in 1926, Golgi continued to think, quite erroneously, that nerve cells are not selfcontained units. For his part, Cajal later wrote of the shared Nobel Prize, "What a cruel irony of fate to pair, like Siamese twins, united by the shoulders, scientific adversaries of such contrasting character."
This disagreement reveals several interesting things about the sociology of science that I was to observe repeatedly during my own career. To begin with, there are scientists, like Golgi, who are very strong technically but who do not necessarily have the deepest insights into the biological questions they are studying. Second, even the best scientists can disagree with one another, especially in the early stages of discovery.
Occasionally, disputes that start out as disagreements about science take on a personal, almost vindictive quality, as they did with Golgi. Such disputes reveal that the qualities that characterize competition— ambition, pride, and vindictiveness—are just as evident among scientists as are acts of generosity and sharing. The reason for this is clear. The aim of science is to discover new truths about the world, and discovery means priority, being there first. As Alan Hodgkin, the formulator of the ionic hypothesis, wrote in an autobiographical essay, "If pure scientists were motivated by curiosity alone, they should be delighted when someone else solves the problem they are working on—but that is not the usual reaction." Recognition by their peers and esteem come only to those who have made original contributions to the common stock of knowledge. This caused Darwin to point out that his "love of natural science . . . has been much aided by the ambition to be esteemed by my fellow scientists."
Finally, great controversies often originate when available methodologies are insufficient to provide an unambiguous answer to a key question. It was not until 1955 that Cajal's intuitions were borne out conclusively. Sanford Palay and George Palade at the Rockefeller Institute used the electron microscope to demonstrate that in the vast majority of cases, a slight space—the synaptic cleft—separates the presynaptic terminal of one cell from the dendrite of another cell. Those new images also revealed that the synapse is asymmetrical, and that the machinery for releasing chemical transmitters, discovered much later, is located only in the presynaptic cell. This explains why information in a neural circuit flows in just one direction.
PHYSIOLOGISTS WERE QUICK TO SEE THE IMPORTANCE OF
Cajal's contributions. Charles Sherrington (figure 4-7) became one of Cajal's greatest supporters and invited him to England in 1894 to give the Croonian Lecture to the Royal Society in London, one of the most distinguished honors Great Britain can bestow on a biologist. In his memorial to Cajal in 1949, Sherrington wrote:
Is it too much to say of him that he is the greatest anatomist the nervous system has ever known? The subject had long been a favorite with some of the best investigators, previous to Cajal there were discoveries, discoveries which often left the physician more mystified than before, adding mystification without enlightenment. Cajal made it possible even for a tyro to recognize at a glance the direction taken by the nerve-current in the living cell, and in a whole chain of nerve cells.
He solved at a stroke the great question of the direction of the nerve-currents in their travel through brain and spinal cord. He showed, for instance, that each nerve-path is always a line of one-way traffic only, and that the direction of that traffic is at all times irreversibly the same.
In his own influential book, The Integrative Action of the Nervous System, Sherrington built on Cajal's findings about the structure of nerve cells and succeeded in linking structure to physiology and to behavior.

4-7 Charles Sherrington (1857-1952) studied the neural basis of reflex behavior. He discovered that neurons can be inhibited as well as excited and that integration of these signals determines the actions of the nervous system. (Reprinted from The Integrative Action of the Nervous System, Cambridge University Press, 1947.)
He did this by examining the spinal cord of cats. The spinal cord receives and processes sensory information from the skin, joints, and muscles of the limbs and trunk. It contains within itself much of the basic neuronal machinery for controlling the movement of the limbs and the trunk, including the movements involved in walking and running. Trying to understand simple neural circuits, Sherrington studied two reflex behaviors—the cat's equivalent of the human knee jerk and the withdrawal response of the cat's paw when exposed to a stimulus that causes an unpleasant sensation. Such innate reflexes require no learning. Moreover, they are intrinsic to the spinal cord and do not require that messages be sent to the brain. Instead, they are elicited instantly by an appropriate stimulus, such as a tap on the knee or exposure of the paw to a shock or a hot surface.
In the course of his research on reflexes, Sherrington discovered something that Cajal could not have anticipated from anatomical stud-
ies alone—namely, that not all nervous action is excitatory—that is, not all nerve cells use their presynaptic terminals to stimulate the next receiving cells in line to transmit information onward. Some cells are inhibitory; they use their terminals to stop the receiving cells from relaying information. Sherrington made this discovery while studying how different reflexes are coordinated to yield a coherent behavioral response. He found that when a particular site is stimulated so as to elicit a specific reflex response, only that reflex is elicited; other, opposing reflexes are inhibited. Thus a tap on the tendon of the kneecap elicits one reflex action—an extension of the leg, a kick. That tap simultaneously inhibits the opposing reflex action—flexion, the drawing backward of the leg.
Sherrington then explored what was happening to the motor neurons during this coordinated reflex response. He found that when he tapped on the tendon of the kneecap, the motor neurons that extend

the limb (the extensors) were actively excited, while the motor neurons that flex the limbs (the flexors) were actively inhibited. Sherrington called the cells that inhibit the flexors inhibitory neurons. Later work found that almost all inhibitory neurons are interneurons.
Sherrington immediately appreciated the importance of inhibition not only for coordinating reflex responses but also for increasing the stability of a response. Animals are often exposed to stimuli that may elicit contradictory reflexes. Inhibitory neurons bring about a stable, predictable, coordinated response to a particular stimulus by inhibiting all but one of those competing reflexes, a mechanism called reciprocal control. For example, extension of the leg is invariably accompanied by inhibition of flexion, and flexion of the leg is invariably accompanied by inhibition of extension. Through reciprocal control, inhibitory neurons select among competing reflexes and ensure that only one of two or even several possible responses is expressed as behavior.
Integration of reflexes and the decision-making capabilities of the spinal cord and brain derive from the integrative features of individual motor neurons. A motor neuron totals up all the excitatory and inhibitory signals it receives from the other neurons that converge upon it and then carries out an appropriate course of action based on that calculation. If and only if the sum of excitation exceeds that of
inhibition by a critical minimum will the motor neuron signal the target muscle to contract.
Sherrington saw reciprocal control as a general means of coordinating priorities to achieve the singleness of action and purpose required for behavior. His work on the spinal cord revealed principles of neuronal integration that were likely to underlie some of the brain's higher cognitive decision making as well. Each perception and thought we have, each movement we make, is the outcome of a vast multitude of basically similar neural calculations.
Some of the details of the neuron doctrine and its implications for physiology had yet to be established in the mid-1880s, when Freud abandoned his basic research studies of nerve cells and their connections. However, he kept abreast of neurobiology and tried to incorporate some of Cajal's new ideas about neurons in an unpublished manuscript, "Project for a Scientific Psychology," written in late 1895, after he had begun to use psychoanalysis to treat patients and had uncovered the unconscious meaning of dreams. Even though Freud became fully immersed in psychoanalysis, his earlier experimental work had a lasting influence on his thought, and therefore on the evolution of psychoanalytic thought. Robert Holt, a psychologist interested in psychoanalysis, has put it this way:
In many respects Freud seems to have undergone a profound re-orientation as he turned from being a neuroanatomical researcher to a clinical neurologist who experimented with psychotherapy, finally becoming the first psychoanalyst. We would be poor psychologists, however, if we imagined that there was not at least as much continuity as change in this development. Twenty years of passionate investment in the study of the nervous system were not easily tossed aside by Freud's decision to become a psychologist instead and to work with a purely abstract, hypothetical model.
Freud called the period he spent studying nerve cells in simple organisms like crayfish, eels, and primitive fish "the happiest hours of my student life." He left those basic research studies after he met and
fell in love with Martha Bernays, whom he later married. In the nineteenth century, one needed an independent income in order to take on a career in research. In view of his poor financial position, Freud turned instead to the establishment of a medical practice that would earn him sufficient income to support a wife and family. Perhaps if a scientific career could have ensured a living wage then, as it does today Freud would be known as a neuroanatomist and a co-founder of the neuron doctrine, instead of as the father of psychoanalysis.

5
THE NERVE CELL SPEAKS
Had I become a practicing psychoanalyst, I would have spent much of my life listening to patients talk about themselves—about their dreams and waking memories, their conflicts and their desires. This is the introspective method of "talk therapy" that Freud pioneered to arrive at deeper levels of selfunderstanding. By encouraging the free association of thoughts and memories, the psychoanalyst helps patients unpack the unconscious memories, traumas, and impulses that underlie their conscious thoughts and behavior.
In Grundfest's laboratory I soon appreciated that to understand how the brain functions, I would have to learn how to listen to neurons, to interpret the electrical signals that underlie all mental life. Electrical signaling represents the language of mind, the means whereby nerve cells, the building blocks of the brain, communicate with one another over great distances. Listening in on those conversations and recording neuronal activity was, so to speak, objective introspection.
GRUNDFEST WAS A LEADER IN THE BIOLOGY OF SIGNALING.
From him I learned that thinking about the signaling function of nerve cells has proceeded in four distinct phases, reaching from the
eighteenth century to a particularly clear and satisfying resolution in the work of Alan Hodgkin and Andrew Huxley two hundred years later. Throughout, the question of how nerve cells communicate has attracted some of the best brains in science.
The first phase dates to 1791, when Luigi Galvani, a biologist from Bologna, Italy, discovered electrical activity in animals. Galvani left a frog's leg hanging on a copper hook from his iron balcony and found that the interaction of the two dissimilar metals, copper and iron, would occasionally cause the leg to twitch, as if it were animated. Galvani could also cause a frog's leg to twitch by stimulating it with a pulse of electricity. After further study, he proposed that nerve cells and muscle cells are themselves capable of generating a flow of electrical current and that the twitch of muscles is caused by the electricity generated by muscle cells—not by spirits or "vital forces," as was commonly believed at the time.
Galvani's insight and his achievement in bringing nervous activity out of the realm of vital forces and into natural science was elaborated in the nineteenth century by Hermann von Helmholtz, one of the first scientists to bring the rigorous methods of physics to bear on a range of problems in brain science. Helmholtz found that the axons of nerve cells generate electricity not as a by-product of their activity, but as a means of producing messages that are carried along their whole length. These messages are then used to carry sensory information about the outside world into the spinal cord and the brain and to transmit commands for action from the brain and spinal cord to the muscles.
In the course of this work, Helmholtz made an extraordinary experimental measurement that changed thinking about electrical activity in animals. In 1859 he succeeded in capturing the speed at which these electrical messages are conducted and found to his amazement that electricity conducted along a living axon is fundamentally different from the flow of electricity in a copper wire. In a metal wire, an electrical signal is conducted at close to the speed of light (186,000 miles per hour). Despite its speed, however, the strength of the signal deteriorates badly over long distances because it is propagated passively. If an axon relied on passive propagation, a sig-

5-1 Edgar, Lord Adrian (1889-1977) developed methods of recording action potentials, the electrical signals nerve cells use for communication. (Reprinted from Essentials of Neural Science and Behavior, Kandel, Schwartz, and Jessell, McGrawHill, 1995.)
nal from a nerve ending in the skin of your big toe would die out before it reached your brain. Helmholtz found that the axons of nerve cells conduct electricity much more slowly than wires do, and they do so by means of a novel, wavelike action that propagates actively at various speeds up to approximately 90 feet per second! Later studies showed that the electrical signals in nerves, unlike signals in wires, do not decrease in strength as they propagate. Thus, nerves sacrifice speed of conduction for active propagation, which ensures that a signal that arises in your big toe arrives at your spinal cord undiminished in size.
Helmholtz's findings raised a set of questions that would occupy physiology for the next hundred years: What do these propagated signals, later called action potentials, look like and how do they encode information? How can biological tissue generate electrical signals? Specifically, what carries the current for the signals?
THE FORM OF THE SIGNAL AND ITS ROLE IN ENCODING INFORMATION
were addressed in the second phase, which began in the 1920s with Edgar Douglas Adrian's work. Adrian (figure 5-1) developed methods of recording and amplifying the action potentials propagated along the axons of individual sensory neurons on the skin, thereby making the elementary utterances of nerve cells intelligible for the first time. In the process, he made several remarkable discoveries about the action potential and how it leads to what we perceive as a sensation.
To record action potentials, Adrian used a thin piece of metal wire. He placed one end of the wire on the outside surface of the axon of a sensory neuron on the skin and then ran the wire to both an ink writer (so he could look at the shape and pattern made by the action potentials) and a loudspeaker (so he could hear them). Every time
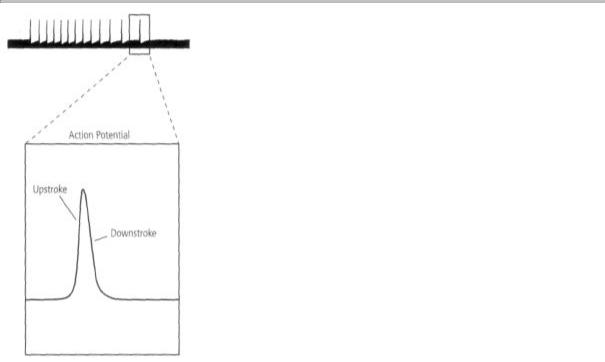
Adrian touched the skin, one or more action potentials were generated. Each time an action potential was generated, he heard a brief bang! bang! bang! over the loudspeaker and saw a brief electrical pulse on the ink writer. The action potential in the sensory neuron lasted only about 1/1000 of a second and had two components: a swift upstroke to a peak, followed by an almost equally rapid downstroke that returned it to the starting point (figure 5-2).
The ink writer and the loudspeaker both told Adrian the same remarkable story: all of the action potentials generated by a single nerve cell are pretty much the same. They are about the same shape and amplitude, regardless of the strength, duration, or location of the stimulus that elicits them. The action potential is thus a constant, all-or-none signal: once the threshold for generating the signal is reached, it is almost always the same, never smaller or larger. The current produced by the action potential is sufficient to excite adjacent regions of the axon, thus causing the action potential to be propagated without failure or flagging along the whole length of the axon at speeds of up to 100 feet per second, pretty much as Helmholtz had earlier found!
The discovery of the all-or-none characteristic of the action potential raised more questions in Adrian's mind: How does a sensory neuron report the intensity of a stimulus—whether a touch is light or heavy, whether a light is bright or dim? How does it signal the dura-
5-2 Edgar Adrian's recordings revealed the characteristics of the action potential. Recordings in single nerve cells showed that action potentials are all-or-none: once the threshold for generating an action potential is reached, the signal is always the same, both in amplitude and shape.
tion of the stimulus? More broadly, how do neurons differentiate one type of sensory information from another, such as touch from pain, light, smell, or sound? How do they differentiate sensory information for perception from motor information for action?
Adrian first addressed the question of intensity. In a landmark finding, he discovered that intensity results from the frequency with which action potentials are emitted. A mild stimulus, such as a gentle touch on the arm, will elicit just two or three action potentials per second, whereas a strong one, such as a pinch or bumping one's elbow, could fire a hundred action potentials per second. Similarly, the duration of a sensation is determined by the length of time over which the action potentials are generated.

Next, he explored how information is conveyed. Do neurons use different electrical codes to tell the brain that they are carrying information about different stimuli, such as pain or light or sound? Adrian found that they did not. There was very little difference among the action potentials produced by neurons in the various sensory systems.
Thus the nature and quality of a sensation—whether visual or tactile, for instance—does not depend upon differences in action potentials.
What, then, accounts for the differences in information carried by neurons? In a word, anatomy. In a clear confirmation of Cajal's principle of connection specificity, Adrian found that the nature of the information conveyed depends on the type of nerve fibers that are activated and the specific brain systems to which those nerve fibers are connected. Each class of sensation is transmitted along specific neural pathways, and the particular kind of information relayed by a neuron depends on the pathway of which it is a part. In a sensory pathway, information is transmitted from the first sensory neuron—a receptor that responds to an environmental stimulus such as touch, pain, or light—to specific and specialized neurons in the spinal cord or in the brain. Thus visual information is different from auditory information because it activates different pathways.
In 1928 Adrian summarized his work in his characteristically vivid style: "all impulses are very much alike, whether the message is destined to arouse the sensation of light, of touch, or of pain; if they are crowded together, the sensation is intense; if they are separated by any interval, the sensation is correspondingly feeble."
Finally, Adrian found that signals sent from motor neurons in the brain to the muscles are virtually identical to signals conveyed by sensory neurons from the skin to the brain: "the motor fibers transmit discharges which are almost an exact counterpart of those in the sensory-fibers. The impulses . . . obey the same all-or-nothing principle." Thus, a rapid train of action potentials down a particular neural pathway causes a movement of our hands rather than a perception of colored lights because that pathway is connected to our fingertips, not to our retinas.
Adrian, like Sherrington, extended Cajal's neuron doctrine, which was based on anatomical observations, into the realm of function. But unlike Golgi and Cajal, who were locked in a bitter rivalry, Sherrington and Adrian were friends who lent each other support. For their discoveries regarding the function of neurons they shared the Nobel Prize in Physiology or Medicine in 1932. On hearing that he would share the prize with Sherrington, Adrian, who was a generation younger, wrote him:
I won't repeat what you must be almost tired of hearing—how much we prize your work and yourself—but I must let you know what acute pleasure it gives me to be associated with you like this. I would not have dreamt of it, and in cold blood I would not have wished it, for your honor should be undivided, but as it is I cannot help rejoicing at my good fortune.
Adrian had listened in on the bang! bang! bang! of neuronal signaling and discovered that the frequency of these electrical impulses represents the intensity of a sensory stimulus, but several questions remained. What lies beneath the nervous system's remarkable ability to conduct electricity in this all-or-none manner? How are electrical signals turned on and off, and what mechanism is responsible for their rapid propagation along an axon?
THE THIRD PHASE IN THE HISTORY OF SIGNALING CONCERNS THE
mechanisms underlying the action potential and begins with the membrane hypothesis, first proposed in 1902 by Julius Bernstein, a student of Helmholtz and one of the most creative and accomplished electrophysiologists of the nineteenth century. Bernstein wanted to know: What mechanisms give rise to these all-or-none impulses? What carries the charge for the action potential?