

МОЛБИОЛ 2014-лекции / ООФ / Л06-Генетическая рекомбинация _1
.pdfЕще одна функция конверсионной системы - контроль над гомеологичной рекомбинацией
•Гомеологическая рекомбинация - кроссинговер между родственными, но дивергировавшими последовательностями ДНК.
•Набор генов, происшедших путем дупликаций и изменений от некоторого генапредка, называется семейством генов.
–Члены одного семейства могут быть расположены рядом и/или разбросаны по разным хромосомам.
–Члены одного семейства могут иметь сходные или идентичные функции, но могут экспрессироваться в разное время или в разных типах клеток.
–К примерам такого рода относятся гены рРНК и гистонов.
•Контроль такой рекомбинации должен препятствовать
–нежелательным перестройкам между членами уже дивергировавших семейств генов и
–генетическим обменам при межвидовой гибридизации.
он обеспечивает стабильность и целостность геномов, с одной стороны, и генетическую изоляцию - с другой.
•Существуют два механизма подавления гомеологичной рекомбинации.
–Основной механизм действует путем подавления едва начавшейся рекомбинации, запрещая миграцию ветвления, с последующим выбрасыванием внедрившейся в гетеродуплекс чужеродной ДНК с помощью хеликазы MutU.
–Второй механизм зависит от белка MutH, но в нем также участвуют MutS и MutL. Детали его пока не выяснены.
31
– У дрожжей и клеток млекопитающих обнаружено участие в этом подавлении
гомологов MutS - белков MSH2 и MSH3.
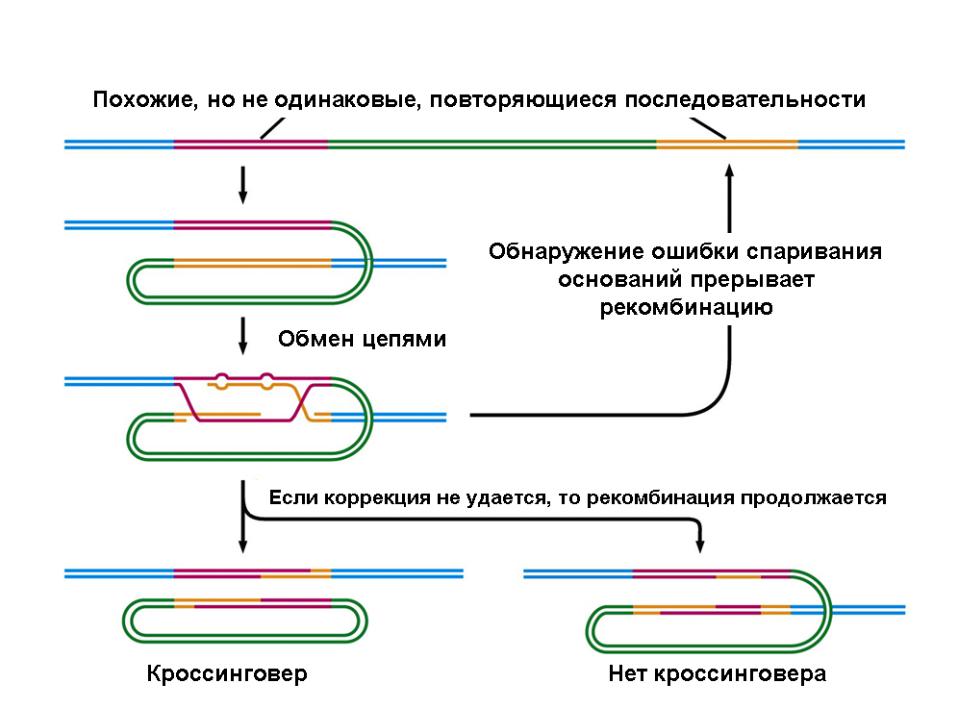
Механизм, предотвращающий дестабилизацию генома
32
Figure 5-67 Molecular Biology of the Cell (© Garland Science 2008)

Основные белки, участвующие в гомологичной рекомбинации
•Процесс гомологичной рекомбинации разделяют на три основные стадии:
–ПРЕСИНАПСИС – подготовка ДНК-субстрата к рекомбинации: введение разрывов, образование участка однонитевой ДНК и т.д.
–СИНАПСИС – нахождение гомологии и обмен цепями между рекомбинирующими ДНК.
–ПОСТСИНАПСИС – миграция и разрешение структуры Холлидея.
По этим стадиям принято классифицировать белки, осуществляющие рекомбинацию
33

Рекомбинация у E. coli
•Клетки E. coli способны к обмену генетической информацией с помощью двух процессов: конъюгации и трансдукции.
Конъюгация
–Контакт между клетками разных половых типов
–Из клетки донора в клетку реципиента передается одна из двух цепей кольцевой ДНК.
–Связь между конъюгирующими бактериальными клетками непрочная и они постепенно расходятся
–В результате расхождения клеток перенос ДНК прекращается.
–Реципиентные клетки получают от донора только часть хромосомной ДНК. полную однонитевую кольцевую, а только ее часть,
–Перенесенная ДНК сразу достраивается до линейного двуцепочечного фрагмента с тупыми концами.
Трансдукция
–Из клеток донора в клетку реципиента с помощью бактериофага переносится двуцепочечный фрагмент бактериальной хромосомы,
но в десятки раз меньшего размера, чем при конъюгации. |
34 |

Основной путь рекомбинации у E. coli - RecBCD
•При конъюгации и трансдукции в реципиентной клетке оказывается линейный дуплекс донорной ДНК с тупыми концами, необходимыйдля рекомбинации с реципиентной хромосомой с участием RecBCD.
•Рекомбинацию инициирует RecBCD-нуклеаза, субъединицы которой кодируются генами recB, recC и recD.:
•Для ферментативной активности ей необходим гидролиз АТФ.
•RecBCD может гидролизовать одно- и двуцепочечную ДНК, притом с обоих концов, то есть работать как экзонуклеаза.
•Имеет хеликазную активность - расплетает дуплекс ДНК, затрачивая на это энергию АТФ.
•Работает как сайт-специфическая эндонуклеаза: расщепляет одноцепочечную ДНК около особой 8-нуклеотидной последовательности 5'-GCTGGTGG-3', называемой Chi-сайтом.
35

Основной путь рекомбинации у E. coli - RecBCD
SSB-белки
Петля ок. 1000 н.
• а. Фермент атакует конец дуплекса и начинает расплетать его. Реакция
асимметрична. RecBCD удерживает 3'-конец таким образом, что образуется одноцепочечная петля примерно в тысячу нуклеотидов
• 5'-цепь выступает в виде свободного хвоста.
• б. Продолжая расплетать дуплекс, RecBCD-нуклеаза продвигает петлю в
3'-цепи вдоль дуплекса. Поскольку после прохождения фермента комплементарные цепи ренатурируют, в 5'-цепи возникает вторая петля.
• в. Двойная петля продвигается вдоль дуплекса до тех пор, пока фермент
не встретит Chi-сайт в 3'-цепи. Фермент подходит к Chi-сайту с 3'-стороны. На расстоянии 4-6 нуклеотидов до него RecBCD разрывает 3'-цепь.
• г. Дальнейшее расплетание дуплекса приводит к вытеснению
рекомбиногенного одноцепочечного 3'-конца, который связывается с белком RecA.
• RecBCD нуклеаза — один из самых ранних белков рекомбинации. Она |
|
готовит субстрат для белка RecA. |
36 |
|
|

Основной путь рекомбинации у E. coli - RecBCD
•д. е. На 3'-конце сначала формируется филамент, затем образуется D-петля (закрытая, так как возникает в кольцевой хромосоме).
•ж. Затем D-петля разрезается с помощью одной из эндонуклеаз E. coli, что приводит к полухиазме Холлидея.
•5'-Концевая часть разорванной цепи ДНК, вероятно, удаляется несколько раньше с помощью экзонуклеазной активности RecBCD.
•з. Далее ДНК-полимераза и ДНК-лигаза должны залечить брешь и разрывы в цепях.
•Такие же события происходят на другом конце
донорной ДНК E.coli, что обеспечивает парность 37
обменов при рекомбинации с хромосомой реципиента.

RecA - главный белок рекомбинации у Е.coli
•В лаборатории А. Кларка в США был выделен и изучен набор мутантов с нарушениями рекомбинации. С помощью этих мутантов, названных rec, были установлены соответствующие гены, а затем выделены белки, участвующие в рекомбинациии.
•На одинарную цепь на 3‘-конце ДНК воздействует несколько белков, которые направляют ее инвазию в дуплекс гомологичной ДНК.
Основным из них является белок RecA (у Е.coli) или его гомолог Rad51 (у эукариот).
•RecA - эволюционно консервативный небольшой белок (около 38 кДа), способный к кооперативному (его молекулы собираются на ДНК по принципу «конец-в-конец») связыванию с любой формой одноцепочечной ДНК.
•Для активации мономера необходимы АТФ и однонитевая ДНК.
•Участвует не только в рекомбинации, но и в репарации ДНК.
•Основное назначение белка RecA - приводить во взаимодействие одноцепочечную ДНК с гомологичным дуплексом.
•Действует кооперативно с белком SSB и ДНК-гиразой;
•Является ДНК-зависимой АТФазой (одна молекула АТР расщепляется до
АDР на 5-10 нуклеотидов ДНК). |
38 |
|

RecA - главный белок рекомбинации у Е.coli
•Свойства белка RecA определяется наличием у него двух сайтов связывания с ДНК.
•Первый сайт используется для первичного связывания с ДНК.
–В присутствии АТФ белок взаимодействует в этом сайте с одноцепочечной ДНК (ssDNA), образуя вокруг ДНК правозакрученную белковую спираль.
–Возникает нитевидное образование - RecA-ДНК-филамент.
•В первых этапах формирования филамента участвует белок SSB, который распрямляет цепь ДНК и позволяет белку RecA строить филамент, но затем SSB вытесняется белком RecA.
•В условиях in vivo в сборке филамента участвуют и другие дополнительные белки.
•Второй сайт RecA используется для связывания RecA-ДНК- филамента со второй молекулой - «голой» двуспиральной ДНК (АТФ-независимая обратимая стадия). Связывание слабое – имеют место лишь кратковременные контакты (соударения). Эти контакты становятся прочными только после нахождения гомологичных последовательностей, когда происходит спаривание в гетеродуплекс.
39

Сборка RecA-филамента
•Сборка филамента происходит в направлении 5’-3’ относительно цепи ДНК, вокруг которой он строится.
•Цепь ДНК располагается вдоль внутренней оси филамента.
•Если однонитевая ДНК находится в составе двунитевой ДНК (хвост или брешь), то формирующийся на ней филамент может перейти на дуплекс.
•Внутри филамента двунитевая ДНК вытянута в 1,5 раза по сравнению с В- формой ДНК.
•В филаменте двуцепочечная ДНК изменяет свою конформацию. В отличие от обычной В-формы ДНК (где шаг спирали составляет 10,4 п.н.) на один виток спирали ДНК в филаменте приходятся 18,6 п.н., в результате чего ДНК оказывается растянутой в 1,5 раза.
•Считается, что такое растяжение дуплекса необходимо для его последующего взаимодействия с гомологичной одноцепочечной ДНК.
• Формирование филамента завершает подготовительную, |
40 |
|
пресинаптическую стадию кроссинговера. |
||
|