

МОЛБИОЛ 2014-лекции / ООФ / Л06-Генетическая рекомбинация _1
.pdfГенетическая рекомбинация (1)
гомологичная рекомбинация
Лекция 6
2014 год
«Структура Холлидея» — промежуточный этап обмена участками гомологичных хромосом

Генетическая рекомбинация
•Генетическая рекомбинация - реорганизация генетического материала посредством разрывов, обмена участками и воссоединения молекул ДНК
•Генетическая рекомбинация является одним из основных источников наследственной изменчивости у всех живых организмов. Это определяет ее важную роль как в эволюции, так и в онтогенетической изменчивости.
•Генетическая рекомбинация участвует в репарации двунитевых разрывов ДНК.
•Для рекомбинации необходим физический контакт между рекомбинирующими участками ДНК – синапсис.
Типы генетической рекомбинации:
Исходя из молекулярных механизмов, генетического контроля и природы синапсиса, все известные рекомбинационные процессы можно подразделить на следующие типы:
«Законная" (общая, или гомологичная)
–обмен гомологичными (подобными или идентичными) участками молекул ДНК;
«Незаконная" (негомологичная) 2
– обмен негомологичными участками ДНК

Типы генетической рекомбинации
•Сайт-специфическая рекомбинация (site-specific recombination)
•Консервативная сайт-специфическая рекомбинация
–обычно происходит между определенными нуклеотидными последовательностями ДНК в пределах очень коротких участков гомологии (15—30 п.о.)
•широко распространена у прокариот и низших эукариот
•интеграция ДНК умеренных фагов в хромосомы бактерий
•инверсия отдельных участков ДНК в хромосомах бактерий и бактериофагов и другие процессы, играющие важную роль в циклах развития фагов и бактерий.
•перестройки в нуклеотидных последовательностях ДНК, кодирующих иммуноглобулины
•Сайт-неспецифическая рекомбинация
•Транспозиция
–перемещение подвижных генетических элементов
3

Гомологичная рекомбинация
•Мейотическая рекомбинация
–У эукариот наиболее типичен обмен участками гомологичных хромосом в мейозе. Этот обмен может происходить между плотно конъюгированными хромосомами на ранних стадиях развития яйца или сперматозоида.
•Митотическая рекомбинация
–Реже гомологичная рекомбинация осуществляется при митозе. В основном направлена на репарацию разрывов в ДНК
–Может сопровождаться нежелательными последствиями (например, возникновением мозаицизма - наличием в тканях генетически различающихся клеток).
–Может быть вызвана рентгеновским облучением клеток на стадии G2 клеточного цикла. Индуцированная митотическая рекомбинация - удобный метод изучения действия генов в процессе развития.
4

Основные функции гомологичной рекомбинации
•Репарация ДНК
–Восстановление разрушенной репликационной вилки
–Точное устранение двухцепочечных разрывов и др. повреждений ДНК
•Обмен генетической информацией между двумя гомологичными хромосомами
–Создаются новые сочетания последовательностей ДНК в каждой из хромосом, что может иметь потенциальную эволюционную выгоду
•Механическая функция - образование и сохранение физической связи между гомологами вплоть до их расхождения в первом делении мейоза
–Если между парой гомологов не произошло ни одного обмена, то они могут неправильно разойтись к полюсам деления. Тогда у одних гамет будет избыточная доза генов, а у других этих генов не будет вовсе. И то, и другое чаще всего ведет к гибели организмов
5
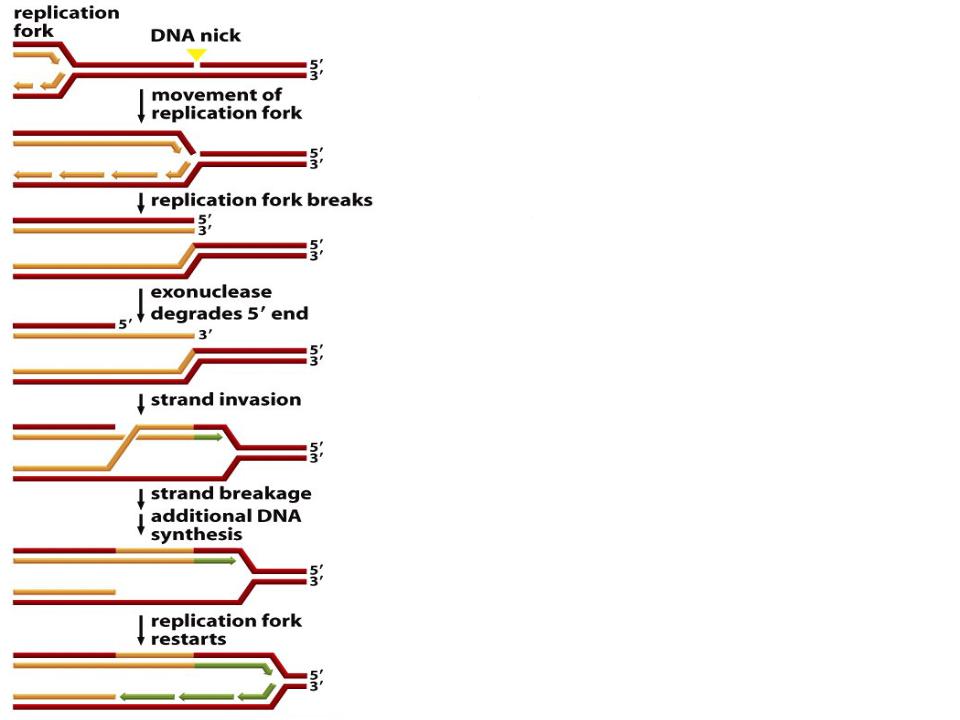
Восстановление разрушенной репликационной вилки при помощи гомологичной рекомбинации
•Подобные «аварии» происходят почти при каждом цикле репликации ДНК
•Когда движущаяся репликационная вилка наталкивается на одноцепочечный разрыв, она разрушается и дает одну разорванную и одну интактную дочерние хромосомы
•Для начала инвазии цепи требуется свободный 3‘-конец, который образуется под действием экзонуклеазы, отщепляющей нуклеотиды с 5‘–конца комплементарной цепи
•После инвазии цепи происходит рекомбинация и возобновление репликации
•Репликационная вилка минует разрыв на одной из цепей, используя в качестве матрицы неповрежденную копию этого6
участка
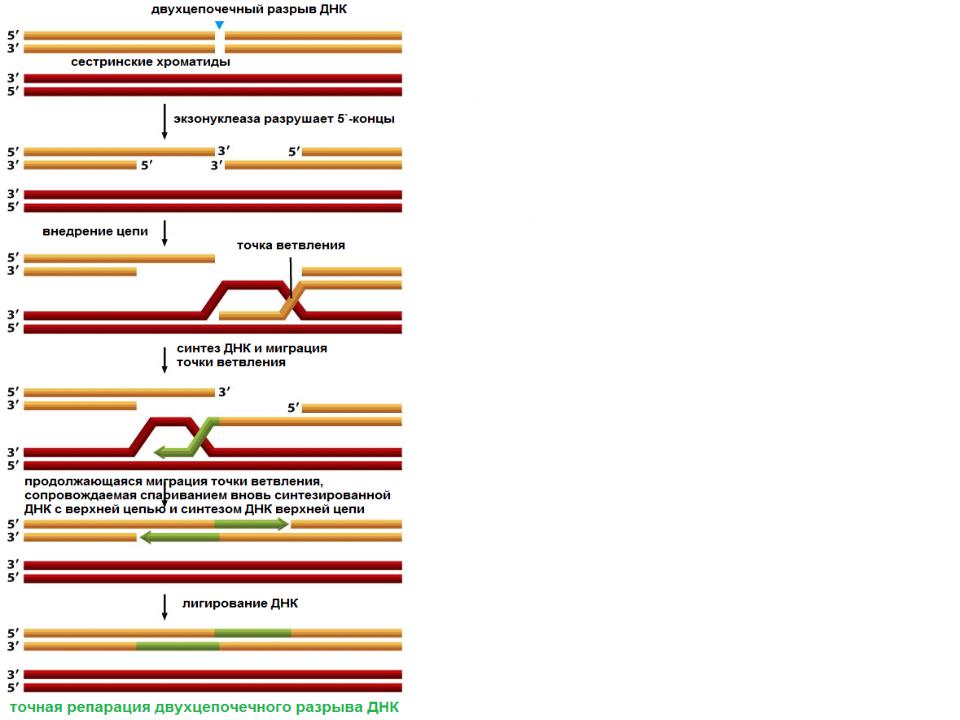
Устранение двухцепочечного разрыва посредством гомологичной рекомбинации
•В большинстве клеток опосредованное гомологичной рекомбинацией устранение двунитевых разрывов происходит только после завершения репликации, и поэтому один из дочерних дуплексов может служить матрицей для восстановления второго
•Гомологичная рекомбинация – наиболее универсальный механизм репарации. Она может быть использована клеткой для устранения других типов повреждений ДНК. Вероятно, это объясняет консерватизм
механизмов и белков, участвующих7
в рекомбинации

Лимитирование использования гомологичной рекомбинации для репарации ДНК
•В ряде случаев гомологичная рекомбинация может представлять опасность для клетки, т.к. в случае неправильного воссоединения
хромосом, гены могут быть повреждены, что является причиной заболеваний, известных как «геномные нарушения»
–Последовательность ДНК в одном гомологе может стать функционально неактивной при ее ремонте с использованием другого гомолога. Такая потеря гетерозиготности часто является решающим моментом при развитии злокачественных опухолей.
–Потенциально возможна основанная на рекомбинации «репарация» неповрежденной ДНК
•Как слишком большой, так и слишком малый уровень гомологичной рекомбинации у людей могут приводить к раку. Первый через потерю гетерозиготности, а второй через увеличение частоты мутаций, вызванных неэффективностью репарации.
•Клетки имеют специальные механизмы контроля за рекомбинацией.
–Сшиваются только очень похожие (высоко гомологичные) последовательности аллелей.
–Ферменты, катализирующие рекомбинационную репарацию, у эукариот вырабатываются на высоком уровне и рассредоточены по всему ядру. При повреждении ДНК они быстро направляются к участкам повреждений и образуют «ремонтные фабрики».
–В этом процессе участвует ряд вспомогательных белков. В частности белковые продукты генов Brca1 и Brca2 (мутации этих генов приводят к повышению вероятности рака молочной железы8).
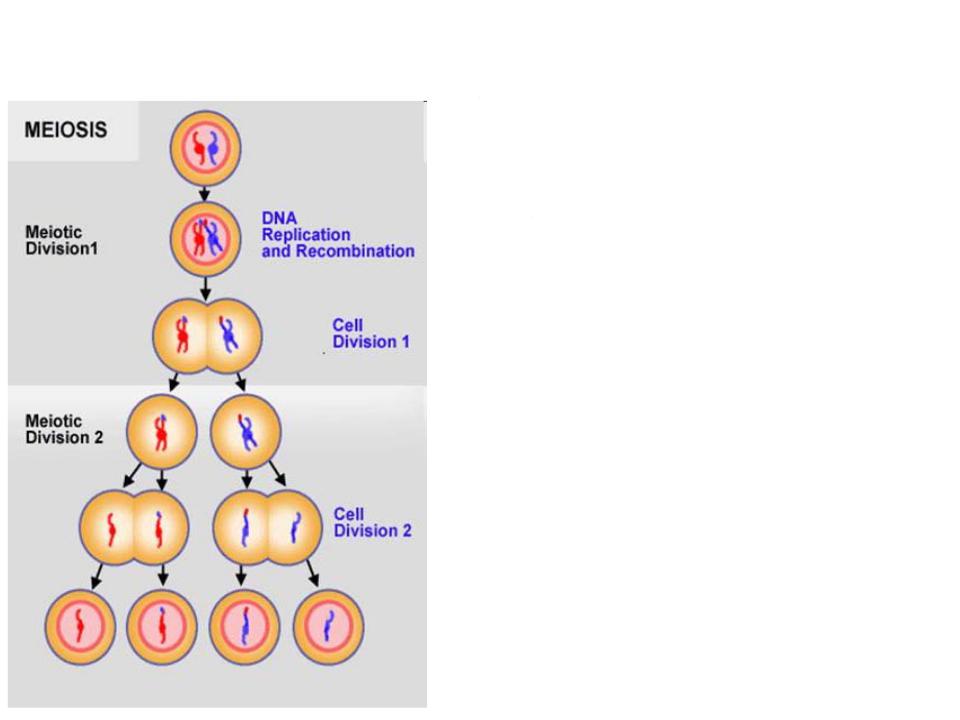
Мейотическая или половая рекомбинация: кроссинговер в профазе первого деления мейоза
•Кроссинговер - процесс обмена участками гомологичных хромосом
•Ядро мейотической клетки в профазе I содержит по четыре гомологичных хроматиды, но в каждом отдельном акте кроссинговера участвуют только две из них.
•Обязательным условием для осуществления кроссинговера является конъюгация хромосом и образование синаптонемального комплекса.
•Конъюгация - соединение гомологичных хромосом
•Однако конъюгация не обязательно влечет за собой кроссинговер
9
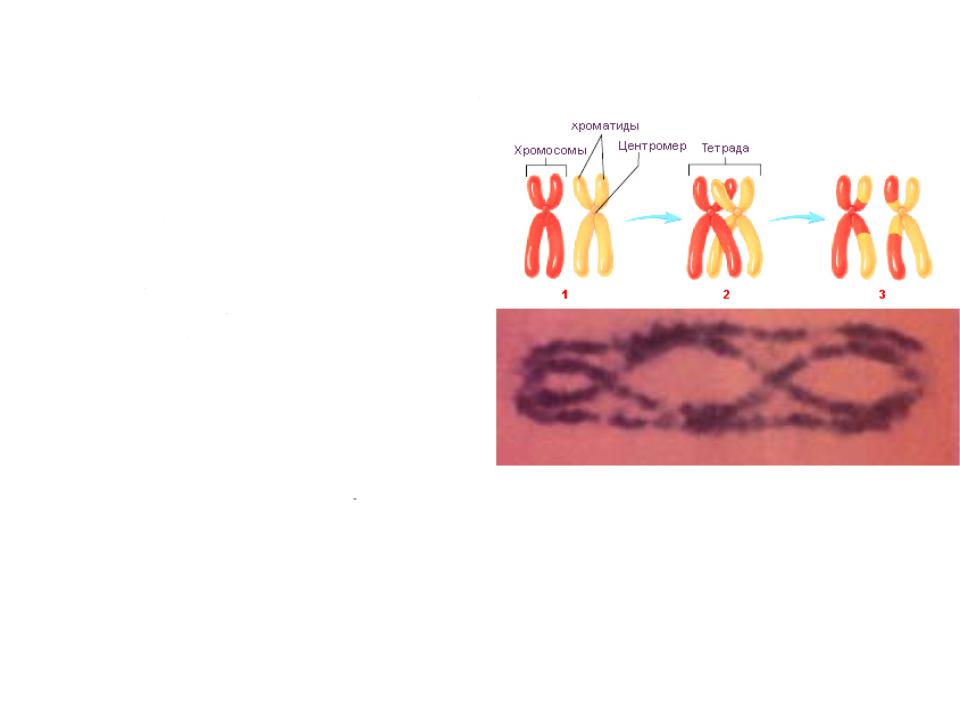
При конъюгации гомологичных хромосом в мейозе между ними происходит кроссинговер
•В биваленте из 4 хроматид в кроссинговере участвуют две, то есть процесс идет на хроматидном уровне, а не на хромосомном
•Объединенные синаптонемальным комплексом в бивалент гомологичные хромосомы, а точнее сестринские хроматиды могут перекрещиваться и обмениваться участками, что приводит к образованию хиазм.
Молекула ДНК состоит из 2 параллельных цепей нуклеотидов; в состав 1 хроматиды входит 1 молекула ДНК; каждая хромосома (вплоть до анафазы II) состоит из 2 хроматид. Соответственно, каждая пара гомологичных хромосом состоит из 2 хромосом, 4 хроматид, 4 молекул ДНК, 8 нуклеотидных цепочек. Рекомбинация происходит между хроматидами отцовской и материнской хромосом.
Образование хиазмы в результате кроссинговера
•Для осуществления кроссинговера необходимо возникновение разрывов в ДНК. Согласно концепции Холлидея рекомбинация осуществляется при согласованном внесении разрывов и воссоединении цепей двух
спиралей ДНК с образованием протяженных гетеродуплексных областей
•В результате возникают качественно новые хромосомы, содержащие
участки (гены) как материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет с новым сочетанием аллелей, 10 получили название кроссоверных или рекомбинантных..