

порок формируется на 4-й неделе эмбриогенеза, в момент закладки переднего мозга. Как и предыдущий, он несовместим с жизнью. Часто встречается у мертворожденных при различных хромосомных и генных синдромах.
Нарушения дифференцировки коры — агирия (отсутствие извилин) и олигогирия с пахигирией (малое количество утолщенных извилин) — сопровождаются упрощением гистологического строения коры. У детей с такими пороками выявляются грубая олигофрения и нарушение многих рефлексов. Большинство детей умирают в течение первого года жизни.
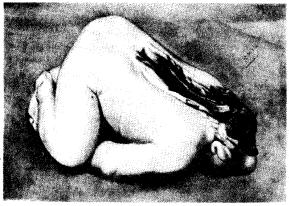
Рис. 14.41. Рахисхиз (платиневрия)
14.6.2. Эндокринная система
Эндокринная система наряду с нервной является ведущим аппаратом интеграции многоклеточного организма, обеспечивая гуморальную регуляцию функций органов. Эта регуляция осуществляется гормонами — биологически активными веществами разной химической природы, выделяемыми железами внутренней секреции. Действие гормонов строго специфично: разные гормоны действуют на разные органы, вызывая определенные изменения их функционирования. Железы внутренней секреции не имеют протоков и выделяют гормоны непосредственно в кровь, что облегчает их транспорт к органам-мишеням. Клетки органов-мишеней на своих мембранах имеют специфические рецепторы, с которыми связываются гормоны, вызывая определенные изменения их метаболизма. Гуморальная регуляция эволюционно возникла значительно раньше нервной потому, что она более проста и не требует развития таких сложных структур, как нервная система.
Обсуждая вопрос о филогенезе эндокринной системы хордовых, необходимо рассмотреть происхождение и эволюцию как гормонов, так и самих желез внутренней секреции.
14.6.2.1. Гормоны
Химическая структура гормонов многообразна. Это могут быть белки
144
(инсулин, пептидные нейрогормоны), стероиды (половые гормоны), продукты метаболизма отдельных аминокислот (тироксин, адреналин). Они могут иметь и другое строение. Это свидетельствует о том, что гормоны разнообразны и по происхождению. Главное, что их объединяет, — способность специфически изменять клеточный метаболизм при контакте с цитоплазматической мембраной.
Предполагают, что уже древние одноклеточные организмы использовали биологически активные вещества для межклеточных коммуникаций. Некоторые вещества такого рода, обладающие регуляторными функциями, могли действовать как на отдельные клетки простейших, так позже и на клетки многоклеточных организмов. Впоследствии они и стали выполнять функции гормонов. Интересно, что в прогрессивной эволюции гуморальной регуляции структура самих гормонов может и не меняться. Доказательством этого является обнаружение таких известных гормонов, как адреналин, норадреналин и некоторые другие, в клетках простейших
инизших растений, где они выполняют функции регуляторов клеточного деления, движения ресничек и вакуолей.
Вэмбриогенезе многоклеточных ряд гормонов выявляется уже в первые часы
идни развития. В процессе дробления они регулируют течение клеточного цикла. Позже — перемещения клеток и образование межклеточных контактов, действуя либо внутри клеток, их продуцирующих, либо на близлежащие клетки. Гормоны приобретают свойства дистантных регуляторов в филогенезе только у трехслойных животных, а в онтогенезе многоклеточных — соответственно на стадии первичного органогенеза.
При неизменности химической структуры функции гормонов могут нередко изменяться. Так, гормон пролактин, выделяющийся у млекопитающих и человека гипофизом и регулирующий секрецию молока млечными железами, обнаружен также у рыб, земноводных и птиц. У первых он регулирует выделение кожными железами слизи, которой питаются мальки, у вторых — образование оболочек икринок в яйцеводах, у третьих — некоторые элементы брачного поведения, а также выделение у кормящих родителей «зобного молочка». Как видно из этого примера, эволюция действия гормонов может быть канализированной выполнением одной и той же функции в самом широком смысле. Действительно, все перечисленные функции пролактина у животных, находящихся на разных уровнях организации, имеют отношение к обеспечению успешности размножения.
Функции других гормонов, например адреналина, могут в филогенетическом ряду тех же позвоночных практически не меняться, обеспечивая у всех регуляцию в первую очередь энергетического обмена.
При этом часто гормоны продолжают выполнять те функции, которые первично проявлялись еще у одноклеточных и низших многоклеточных животных. Так, адреналин в организме млекопитающих и человека замедляет прохождение клеток по клеточному циклу и вступление их в митоз. Из главной эта функция у высокоорганизованных животных превратилась во второстепенную. Таким образом, на филогенез гормонов, так же как и на эволюцию органов, распространяются основные закономерности макроэволюции (см. гл. 13).
145
14.6.2.2. Железы внутренней секреции
Железы внутренней секреции, как и гормоны, выделяемые ими, имеют разное происхождение, что важно для изучения их эволюции.
Некоторые эндокринные железы связаны по происхождению с эпителиальной выстилкой глотки. К ним относятся щитовидная и паращитовидная железы. Эпифиз развивается как вырост мозга; гипофиз, надпочечники и поджелудочная железа имеют сложное происхождение.
Среди хордовых только у бесчерепных эндокринная система существует в виде отдельных клеток и клеточных комплексов, которые находятся в разных отделах тела, объединенных друг с другом за счет гуморального взаимодействия. У позвоночных в основании промежуточного мозга развивается гипоталамус — нейросекреторное образование, осуществляющее связь между двумя системами интеграции организма в единое целое: нервной и эндокринной. Вместе с гипофизом гипоталамус образует единую гипоталамо-гипофизарную систему.
Эволюционным предшественником гипоталамуса является так называемый инфундибулярный вырост ланцетника, состоящий из нейросекреторных клеток и находящийся на вентральной стороне переднего конца нервной трубки.
Начиная с рыб гипоталамус дифференцируется на многочисленные ядра, клетки которых с помощью отростков контактируют как с нейронами мозга, так и с клетками гипофиза. Нейросекреторные клетки гипоталамуса выделяют две основные группы гормонов: пептидные и моноаминовые.
Первые — гормоны, влияющие на функции внутренних органов — вазопрессин, регулирующий артериальное давление, окситоцин, действующий на мускулатуру матки, и др. Вторая группа гормонов (дофамин, норадреналин, серотонин) регулирует деятельность передней доли гипофиза. Под их действием стимулируется или подавляется секреция гормонов соответствующими гипофизарными клетками.
Подробнее остановимся на эволюции гипофиза. Эта железа состоит из трех долей: передней (аденогипофиза), средней (промежуточной) и задней (нейрогипофиза). Гипофиз соединен с гипоталамусом его выростом — воронкой, через которую проходят отростки нейронов гипоталамуса и кровеносные сосуды, обеспечивающие их гуморальное взаимодействие.
Доли гипофиза имеют разное происхождение. Передняя доля развивается из выпячивания эктодермального эпителия крыши ротовой полости, так называемого кармана Ратке, который растет в сторону промежуточного мозга. Задняя доля развивается из задней части воронки. Клетки, входящие в ее состав, по происхождению являются глиальными. Промежуточная доля — производная от передней (рис. 14.42).
146
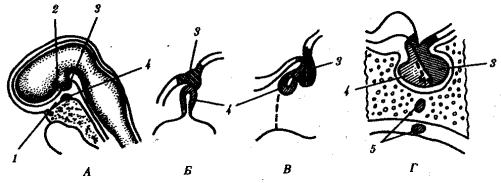
Рис. 14.42. Развитие гипофиза у человека. А—сагиттальный срез 4-недельного зародыша; Б—объединение зачатков гипофиза из стенки глотки и основания мозга; В—формирование трех долей гипофиза на 8-й неделе развития; Г—сформированный гипофиз:
1—крыша ротовой полости, 2—основание мозга, 3—выпячивание основания мозга (задняя доля гипофиза), 4—карман Ратке (промежуточная и передняя доли гипофиза), 5—аномалия развития (добавочные дольки передней доли в стенке глотки
ивнутри клиновидной кости, оставшиеся в области выпячивания кармана Ратке)
Ухрящевых рыб во взрослом состоянии сохраняется первоначальная связь передней доли гипофиза с эпителием ротовой полости. За счет ее задней части формируется также средняя доля. Обе доли вырабатывают гонадотропные гормоны. У костных рыб и личинок земноводных имеются передняя и промежуточная доли, а у взрослых амфибий, переходящих к наземному существованию, появляется также задняя, регулирующая водный обмен. Средняя доля у них перестает выделять гонадотропный гормон, но вырабатывает пролактин.
В связи с наземным образом жизни пресмыкающихся и млекопитающих у них наиболее прогрессивно развивается задняя доля гипофиза, что связано с интенсификацией водного обмена.
Передняя доля вырабатывает соматотропный гормон (гормон роста) и ряд гормонов, регулирующих функции других желез внутренней секреции, а средняя — пролактин и некоторые другие. При этом усиливается дифференцировка и интенсифицируются функции ядер гипоталамуса, находящихся в функциональной связи со всеми долями гипофиза.
Учеловека в эмбриогенезе развитие гипофиза соответствует основным этапам его эволюции. Очень часто, в 30—40%, у нормальных людей под слизистой оболочкой крыши глотки, в основании клиновидной кости, обнаруживается группа клеток длиной 5—6 мм и шириной 0,5—1 мм, по структуре и функциям соответствующая передней доле гипофиза. Это результат нарушения перемещения клеток при закладке гипофиза в эктодерме ротовой полости в области турецкого седла. Эту 139 аномалию называют эктопией аденогипофиза, она не сопровождается
патологическими проявлениями. Более опасно сохранение полости в области
147
кармана Ратке — киста кармана Ратке. Она располагается между передней и промежуточной долями, содержит слизь и в ряде случаев имеет тенденцию к росту и даже к переходу в злокачественное новообразование. Растущие кисты сдавливают гипофиз и вызывают его гипофункцию. Больные нуждаются в оперативном вмешательстве.
Щитовидная железа, гормон которой — тироксин — регулирует энергетический обмен, среди хордовых как компактный орган впервые появляется у рыб. Однако уже у ланцетника отдельные тироксин-синтезирующие клетки обнаруживаются в желобке на вентральной стороне глотки. Щитовидная железа рыб закладывается также в виде желобка на вентральной стороне глотки между 1-й и 2-й жаберными щелями в области зачатка основания языка. Позже этот клеточный материал погружается под слизистую оболочку и формирует фолликулы, характерные для щитовидной железы. У других позвоночных железа закладывается так же, как у рыб, но затем она перемещается в область подъязычной кости (у земноводных) или в шейную область (у пресмыкающихся и млекопитающих).
У человека в эмбриогенезе щитовидной железы происходит рекапитуляция предковых состояний. Гетеротопия ее осуществляется посредством миграции клеток в виде тяжа, полого внутри, называющегося щитоязычным протоком — canalis thyreoglossus. При нормальном развитии этот проток полностью редуцируется, рудиментом его является слепое отверстие foramen caecum в корне языка.
Свидетельством эмбрионального перемещения железы является и расположение верхней щитовидной артерии a. thyreoidea superior, которая, начинаясь от общей сонной артерии, резко поворачивает вниз и входит в ее ткань. На ранних этапах эмбрионального развития эта артерия направляется вверх к зачатку щитовидной железы, а затем меняет свое направление вместе с его перемещением книзу.
Персистирование участков подъязычного протока в постнатальном периоде сопровождается накоплением в них жидкости и образованием срединных кист шеи, которые могут располагаться в любом месте от корня языка до верхней границы щитовидного хряща. Иногда, кисты загнаиваются и прорываются с образованием срединных свищей шеи. Не менее известны такие пороки развития, как эктопия щитовидной железы, которая может развиться в результате нарушения клеточной миграции (рис. 14.43).
Из эпителия глотки в области III-V жаберных карманов у позвоночных развиваются мелкие эндокринные образования, связанные по месту окончательного положения с щитовидной железой. Это паращитовидные железы и ультимобранхиальные тельца. Первые, выделяя гормон паратиреоидин,
повышающий содержание ионов кальция в крови и уменьшающий их количество в костях, развиваются как самостоятельные железы только у наземных позвоночных, а у земноводных — лишь после метаморфоза. Клетки вторых выделяют кальцитонин, являющийся антагонистом паратиреоидина. Таким образом, паращитовидные железы и ультимобранхиальные тельца являются регуляторами кальциевого обмена. Значение их наиболее велико у наземных позвоночных. В филогенетическом ряду
148