

способами. Так, грызуны и копытные млекопитающие питаются одинаковой растительной пищей, но у первых наиболее выраженные адаптации к растительноядности проявляются в строении зубов и морфофизиологии слюнных желез, в то время как вторые на фоне примитивной зубной системы имеют высокоспециализированные желудок и кишечник. Явления гетеробатмии и, следовательно, компенсации функций имеют огромное эволюционное значение в связи с тем, что в организме, даже вступившем на путь узкой специализации, всегда остаются органы и системы относительно мало специализированные, которые при меняющихся условиях могут еще прогрессивно развиваться, раскрывая перед такими филогенетическими группами новые адаптивные возможности.
13.5. СОВРЕМЕННАЯ СИСТЕМА ОРГАНИЧЕСКОГО МИРА
Все живые организмы подразделяются на неклеточных (вирусы) и клеточных (все остальные). Несмотря на то что филогенетические взаимоотношения между ними неясны, вирусы как облигатно-парази-тические формы (см. § 18.3), возможно, возникли от более высокоорганизованных организмов за счет упрощения в процессе адаптации к паразитизму. В то же время не исключена возможность существования вирусов как фрагментов нуклеиновых кислот еще на предбиологическом этапе эволюции и приобретения ими основных свойств живых организмов позже, при попадании в клетки. Клеточные организмы подразделяются на про- и эукариот. Эукариоты, вероятно, произошли от прокариот (см. § 1.5).
13.5.1. Типы питания и основные группы живых организмов в природе
Древние эукариоты, будучи одноклеточными, по характеру питания специализировались на группы организмов, активно добывающих пищу путем ее поиска и захвата, и формы, добывающие продукты питания за счет их всасывания из среды. Первый способ питания называют голозойным, второй — голофитным.
Прогрессивная эволюция первого способа питания сопровождает возникновение царства Животные Animalia и весь их последующий филогенез. Поэтому главными чертами животного организма являются способность к перемещению, активному захвату и переработке пищи. Второй способ питания, наоборот, предусматривает приобретение клетками дополнительных защитных оболочек, усложняющих их захват и переваривание и, следовательно, неподвижность. Клетки части организмов, эволюционирующих по этому пути, приобрели клеточную стенку из полисахарида хитина, позволяющую всасывание высокомолекулярных органических веществ. Такие организмы перешли к сапрофитному питанию, т. е. к потреблению разлагающихся органических веществ, и дали начало филогенезу царства Грибы Fungi.
Клеточная стенка других организмов построена из целлюлозы, позволяющей всасывать из среды лишь воду, углекислый газ и неорганические ионы. Приобретение ими способности к фотосинтезу явилось ключевым моментом в
81
прогрессивной эволюции царства Растения Plantae.
Подробнее остановимся на филогенезе царства Животные. Важнейшим прогрессивным событием в эволюции животных является возникновение многоклеточности.
13.5.2. Происхождение многоклеточных животных
Прежде всего необходимо определить понятие многоклеточное животное. Важнейшими чертами многоклеточности являются следующие: 1) тело животного состоит из большого количества клеток; 2) клетки дифференцированы на половые и соматические, а последние различаются также по структуре и функциям; 3) клетки расположены в организме в несколько слоев; 4) клетки интегрированы в целостную систему благодаря существованию жидкостной внутренней среды и нервной системы.
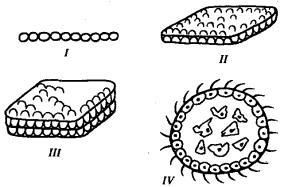
В процессе эволюции первая характеристика многоклеточности достигается наиболее просто: среди простейших уже имеется огромное количество видов колониальных организмов. Остальные черты многоклеточности связаны с возникновением многослойности, обеспечивающей как различные условия существования клеткам, расположенным на поверхности и внутри тела, так и появление внутренней среды, объединяющей их воедино. На рис. 13.10 представлены различные варианты объединения клеток в надклеточные комплексы. Ясно, что только интеграция клеток в шаровидную структуру дает им возможность оказаться в разных условиях, дифференцироваться и взаимодействовать друг с другом.
Рис. 13.10. Варианты объединения клеток в надклеточные образования:
I—нитевидная колония клеток, II—колония в виде однослойного пласта клеток, III— двуслойная колония клеток, IV—сферический многоклеточный организм с клетками разных типов
Родоначальником многоклеточных в настоящее время считают шаровидную колонию жгутиковых, половые клетки которых перемещались в глубь колонии, а
82
соматические первично выполняли как функцию перемещения всей колонии в пространстве, так и пищеварения за счет переваривания фагоцитированных пищевых частиц, захваченных из воды.
Осуществление одной и той же клеткой функций движения и пищеварения малоэффективно. С этим связана последующая специализация клеток в направлении преимущественно пищеварения или обеспечения движения. Результатом является возникновение фагоцитобласта (внутреннего слоя амебовидных клеток, занимающихся пищеварением) и кинобласта (наружного слоя клеток со жгутиками, обеспечивающими движение).
Стойкая дифференцировка соматических клеток по функциям и строению, возникшая первоначально на фоне выделения двух клеточных слоев, явилась ключевым моментом в происхождении многоклеточных. Именно с двуслойностью связано появление жидкой внутренней среды, через которую клетки обмениваются химическими сигналами, а также дальнейшее обособление и специализация части поверхностных клеток в направлении восприятия внешних раздражителей и передача возбуждения на другие клетки, располагающиеся в отдалении от них. Таким образом возникают предпосылки к формированию нервной системы.
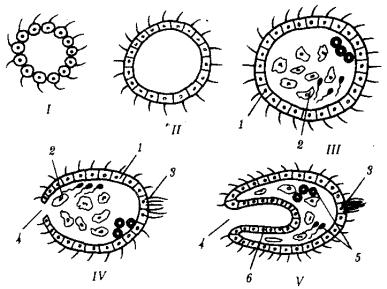
Гипотетический предок многоклеточных животных назван фагоцителлой (рис. 13.11). Он плавал в толще воды за счет биения ресничек кинобласта, а питался, захватывая взвешенные в среде частички пищи и переваривая их клетками фагоцитобласта. На более поздних этапах эволюции происходили многочисленные адаптации потомков фагоцителлы к многообразным условиям существования при оседании их на дно или при перемещении к поверхности, а также при изменении источников питания (захват мелких или крупных, живых или мертвых пищевых частиц).
Рис. 13.11. Этапы происхождения многоклеточности: I, II—сферические колонии жгутиковых, III—V—фагоцителлы разной степени сложности; 1—кинобласт, 2—
83
рыхлый фагоцитобласт, 3—скопление чувствительных клеток на переднем конце тела, 4—ротовое отверстие, 5—половые клетки, 6—эпителизованный фагоцитобласт
Большое значение в эволюции потомков фагоцителлы имели также изменения характера движения: пассивное движение или прикрепленный образ жизни обусловливают лучевой тип симметрии, в то время как активное перемещение в определенном направлении предусматривает формирование двубоковой, или билатеральной, симметрии. В результате возникло огромное многообразие форм многоклеточных животных.
Представления о происхождении многоклеточных, изложенные здесь, являются развитием гипотезы И.И. Мечникова (1877—1880) о том, что многоклеточность возникла на базе шаровидных колоний жгутиковых, часть клеток которых иммигрировала внутрь для переваривания пищевых частиц, в результате чего оформилась дифференцировка на экто- и энтодерму.
13.5.3. Основные этапы прогрессивной эволюции многоклеточных животных
Важным шагом в эволюции многоклеточных животных явилось возникновение третьего зародышевого листка — мезодермы. Мезодерма обеспечивает возможность дифференцировки мышечной, соединительной тканей и скелета, а также многоклеточных половых желез, в которых созревающие гаметы оказываются надежно защищены от неблагоприятных средовых воздействий. Практически все трехслойные животные ведут активно подвижный образ жизни, благодаря чему приобретают билатеральный тип симметрии. Вместе с тем у трехслойных животных с интенсивным обменом веществ, активно перемещающихся с помощью мышц, возникают проблемы с выведением большого количества продуктов диссимиляции из тканей — производных мезодермы, в то время как эктодермальные и энтодермальные клетки выделяют их за счет диффузии соответственно либо наружу, либо в просвет пищеварительной полости. Поэтому именно у трехслойных впервые появляется и прогрессивно эволюционирует
выделительная система.
Следующий значительный этап эволюции животных — возникновение вторичной полости тела, или целома, первоначально функционирующего как гидростатический скелет, а также выполняющего половую и выделительную функции в связи с тем, что продукты диссимиляции и половые клетки попадают в целом и только потом выделяются наружу (см. § 14.5).
84
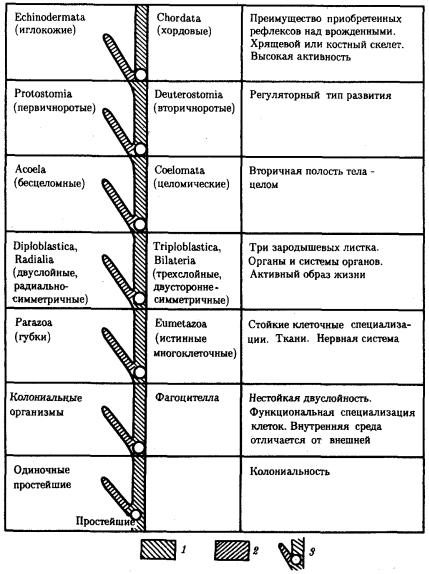
Рис. 13.12. Главные направления эволюции групп в животном царстве: 1—прогрессивное направление, 2—адаптивное направление, 3—узловые моменты в прогрессивной эволюции; каждому узловому моменту соответствует его характеристика, обозначенная в правом столбце
Существенным этапом дальнейшей эволюции многоклеточных является возникновение регуляторного типа эмбрионального развития (см. разд. 8.3.1 и 8.3.2), в результате которого в развивающемся зародыше доминирует целостность морфогенетических процессов над их составляющими. Благодаря этому зародыш развивается относительно автономно в соответствии со своей генетической программой и способен компенсировать даже серьезные повреждения. Организмы, характеризующиеся такими особенностями, относят к группе вторичноротых, в отличие от первичноротых, у которых эмбриональное развитие протекает по мозаичному типу (см. разд. 8.3.1).
Наиболее крупные систематические группировки в царстве Животные называют типами. За период существования жизни на Земле их было не менее 35. К настоящему времени некоторые из них вымерли; сейчас на Земле обитают животные
85