

Рис. 14.16. Болезнь Шпренгеля (пояснение см. в тексте)
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к прямохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохождению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формируются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Они являются фактически анаболиями, возникшими в ходе филогенеза приматов. Поэтому атавистические аномалии скелета, связанные с задержками развития признаков, характерных только для человека, встречаются наиболее часто. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.
14.2.2.Мышечная система
Упредставителей типа Хордовые мускулатура подразделяется по характеру развития и иннервации на соматическую и висцеральную.
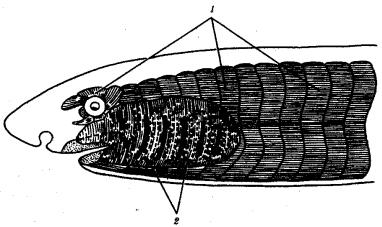
Соматическая мускулатура развивается из миотомов и иннервируется
нервами, волокна которых выходят из спинного мозга в составе брюшных корешков спинномозговых нервов. Висцеральная мускулатура развивается из других участков мезодермы и иннервируется нервами вегетативной нервной системы. Вся соматическая мускулатура поперечно-полосата, а висцеральная может быть как поперечно-полосатой, так и гладкой (рис. 14.17).
Рис. 14.17. Соматическая и висцеральная мускулатура позвоночных: 1—соматическая мускулатура, развивающаяся из миотомов, 2—висцеральная
110
мускулатура жаберной области
14.2.2.1. Висцеральная мускулатура
Наиболее существенные изменения претерпела висцеральная мускулатура, связанная с висцеральными дугами передней части пищеварительной трубки. У низших позвоночных большая часть этой мускулатуры представлена общим сжимателем висцерального аппарата — m. constrictor superficialis, покрывающим всю область жаберных дуг со всех сторон. В области челюстной дуги эта мышца иннервируется тройничным нервом (V), в области подъязычной дуги — лицевым (VII), в области первой жаберной дуги — языкоглоточным (IX), наконец, ее часть, лежащая каудальнее,— блуждающим нервом (X). В связи с этим все производные соответствующих висцеральных дуг и мышц, связанных с ними, иннервируются впоследствии у всех позвоночных перечисленными нервами.
В передней части сжимателя обособляется крупная мускулатурная масса, обслуживающая челюстной аппарат. Позади висцерального аппарата дифференцируется трапециевидная мышца m. trapezius, отдельными пучками прикрепляющаяся к последним жаберным щелям и переднему краю спинного отдела плечевого пояса. Часть поверхностного сжимателя в области подъязычной дуги у пресмыкающихся разрастается, охватывает шею снизу и с боков и образует сжиматель шеи m. sphincter colli. У млекопитающих эта мышца делится на два слоя: глубокий и поверхностный. Глубокий сохраняет прежнее название, а поверхностный называется platysma myoides и располагается подкожно. Эти две мышцы разрастаются на всю область головы и дают начало сложной системе лицевой подкожной мускулатуры, которую у приматов и человека называют мимической. Поэтому вся мимическая мускулатура иннервируется так же, как и мышца, из которой она происходит,— лицевым нервом.
Мышцы собственно жаберного аппарата с утратой жаберного дыхания редуцируются, но отдельные их элементы сохраняются в виде мышц подъязычного аппарата, глотки и гортани. Трапециевидная мышца полностью теряет связь с жаберным аппаратом и становится исключительно мышцей плечевого пояса. Часть ее у млекопитающих отходит от сосцевидного отростка черепа и приклепляется к ключице и грудине, обособляется — грудино-ключично-сосцевидная мышца т. sternocleidomastoideus. Иннервирующая эту мышцу задняя часть блуждающего нерва становится самостоятельным черепно-мозговым нервом XI пары, добавочным — п. accessorius.
Основные этапы филогенеза висцеральной мускулатуры жаберной области рекапитулируют в эмбриогенезе млекопитающих и человека. Знание этих рекапитуляции позволяет объяснить сложность иннервации мышц лица и шеи, объединенных с ними общностью происхождения.
111
14.2.2.2. Соматическая мускулатура
Мускулатура головы. У всех позвоночных животных в процессе эмбриогенеза нижние концы миотомов образуют выросты в вентральном направлений, охватывающие полость тела снаружи и срастающиеся по центральной линии на брюшной стороне. Таким образом, закладки соматической мускулатуры становятся сегментарными не только на дорсальной стороне в связи с сегментацией сомитов, но и на вентральной. В миотомах и в их вентральных отростках идет образование продольных мускульных волокон.
Миотомы, лежащие на головном конце тела, распадаются на мезенхиму и образуют зачатки отдельных мышц. Из первого миотома головы формируются верхняя внутренняя и нижняя прямые и нижняя косая мышцы глаза, иннервируемые глазодвигательнъш нервом п. oculomotorius (III пара). Из второго миотома — верхняя косая мышца, иннервируемая блоковым нервом п. trochlearis (IV пара); а из третьего — наружная прямая мышца, получающая иннервацию от отводящего нерва п. abducens (VI пара).
Задние миотомы головы, образующие мощные вентральные отростки, распространяющиеся вокруг висцеральной мускулатуры глоточной области, образуют подъязычную мускулатуру, которая у рыб слабо дифференцирована, а начиная с земноводных распадается на m. sternohyoideus, т. omohyoideus и т. geniohyoideus. У наземных позвоночных за счет последней мышцы формируется собственная мускулатура языка — m. genioglossus и m. hyoglossus. Вся подъязычная мускулатура иннервируется подъязычным нервом п. hypoglossus, который у амниот становится типичным черепно-мозговым нервом.
Мускулатура туловища и конечностей. У бесчерепных, а также у рыб вся мускулатура туловища состоит из ряда мышечных сегментов, или миомеров, правой и левой сторон, которые вместе образуют так называемые боковые мышцы. Каждый миомер развивается из миотома одного сомита и иннервируется первоначально двигательной ветвью одного спинномозгового нерва. Миомеры отделены друг от друга миосептами — соединительнотканными перегородками. Такой же перегородкой, идущей продольно, бокойая мышца разделена на спинную и брюшную мышцы.
Уже у рыб направление пучков мышечных волокон в миомерах начинает изменяться на разной глубине мышечного слоя. Эта дифференцировка значительно более выражена у наземных позвоночных и приводит у них к постепенному обособлению различных слоев брюшной и спинной мускулатуры. В результате этого возникают сложные группы мышц, от первоначальной четкой метамерии которых остаются только следы в виде глубоких мышц спины и шеи, связывающих друг с другом соседние позвонки. Значение спинной мускулатуры наземных позвоночных снижается в связи с передвижением большинства из них с помощью конечностей, а брюшные мышцы претерпевают смену функций: первоначально принимающие участие в перемещениях тела, они у пресмыкающихся и млекопитающих служат для изменения объема грудной и брюшной полостей в процессе дыхания.
112