

11. Гладкая мышца: значение для организма, функциональная единица, отличия потенциала покоя и потенциала действия от таковых скелетной мышцы, свойства.
Медленные часто ритмические сокращения гладкомышечных стенок внутренних органов (кишечник, желудок, мочеточники, протоки пищеварительных желез) обеспечивает перемещение содержимого этих органов. Тоническое сокращение стенок артерий и артериол поддерживает оптимальный уровень кровяного давления и кровоснабжение органов и тканей.
Гладкие мышцы построены из веретенообразных одноядерных мышечных клеток. Волокна связаны между собой низкоомными щелевыми контактами. Гладкая мышца функционирует как синцитий – функциональное образование, в котором возбуждение способно беспрепятственно передаваться с одной клетки на другую по крайней мере в пределах одного мышечного пучка, являющегося функциональной единицей гладкой мышцы.
ПП обнаруживает постоянные небольшие колебания, обеспечивающие возникновение ПД при достижении КП. Величина ПП меньше, чем в скелетных мышцах.
ПД двух основных типов: пикоподобные и ПД с выраженным плато.
При удалении из омывающего раствора ионов Na возникают полноценные ПД у гладких мышц. Удаление ионов Са ведет у обратимому угнетению ПД.
Свойства гладких мышц
Гладкие мышцы являются тоническими. Их сокращения медленные, длительные, но довольно сильные и не требуют больших энергетических затрат. Расслабление мышечных волокон после их сокращения протекает очень медленно. Гладкие мышцы способны сохранять приданную им растяжением длину без изменения напряжения. Возбудимость гладких мышц низкая. Обычно они возбуждаются действием не одного, а нескольких импульсов. При этом даже относительно редкие раздражения могут вызвать стойкое непрерывное сокращение.
Скорость проведения импульсов и волны сокращения гладких мышц значительно меньше скелетных (от 2 до 15 смс). Работа гладкой мускулатуры регулируется импульсами, поступающими к ней по вегетативным нервам: меняется уровень ее активности, т. е. сила и частота сокращения. Гладкомышечные волокна способны сокращаться благодаря их растяжению без участия нервных воздействий.
12. Рецептор: понятие, функция, классификация рецепторов, свойства и их особенности, механизм возбуждения первичных и вторичных рецепторов, регуляция функции рецепторов.
Рецепторы – структурные элементы клеточной мембраны, воспринимающие действие определенного вида раздражителя и запускающие формирование ответной реакции клетки.
По назначению рецепторы классифицируют на сенсорные, эффекторные и нейронные.
Сенсорные рецепторы – рецепторы, воспринимающие действие адекватного физического или химического раздражителя и запускающие процесс формирования сигнала для посылки в ЦНС или в вегетативный ганглий. Это чувствительные нервные окончания или специальные рецепторные клетки. Восприятие раздражителя осуществляется с помощью преобразования энергии раздражителя в РП, который посредством своего электрического поля обеспечивает возникновение ПД в окончании чувствительного нейрона и посылку сигнала по нервному волокну.
Эффекторные – рецепторы, воспринимающие действие гормона или медиатора и запускающие процесс формирования ответной реакции самой клетки. Это рецепторы мышечных и секреторных клеток.
Нейронные – постсинаптические рецепторы нейронов, воспринимающие действие медиатора и запускающие процесс обработки информации и ее хранения в нейронах, а также посылку сигналов к другим нейронам ЦНС и приказов к рабочим клеткам.
Сенсорные рецепторы разделяют на механорецепторы, хеморецепторы, терморецепторы, фото, ноцирецепторы.
По расположению в организме выделяют экстеро и интерорецепторы, центральные и переферические.
По принадлежности к органам чувств зрительные, слуховые, обонятельные, вкусовые, тактильные.
В зависимости от специфичности моносенсорные и полисенсорные.
По структурно – функциональной организации и электрофизиологическим особенностям разделяют первичные и вторичные. Первичные представляют собой чувствительные окончания дендрита афферентного нейрона. Их тела расположены в спинальных ганглиях или ганглиях черепных нервов. РП генерирует ПД в пределах одной клетки – сенсорного нейрона. К ним относятся обонятельные, тактильные, температурные, болевые и проприорецепторы. Во вторичных рецепторах имеется специальная клетка эпителиального или нейроэктодермального происхождения, синаптически связанная с окончанием дендрита сенсорного нейрона. РП и ПД возникают в разных клетках. РП формируется в рецепторной клетке, а ПД в окончаниях отростков сенсорного нейрона. К ним относятся фоторецепторы, слуховые, вкусовые, вестибулярные.
По скорости адаптации различают быстро и медленно адаптирующиеся, смешанные и адаптирующие со средней скоростью.
Свойства рецепторов.
Высокая возбудимость
Спонтанная активность
Адаптация
Регуляция функции рецепторов осуществляется НС и гуморальными
Факторами. Нервные центры осуществляют эфферентный контроль за работой вестибулярных, слуховых, обонятельных и мышечных рецепторов.
НС может регулировать возбудимость рецепторов изменяя концентрацию гормонов, которые, действуя на белки рецепторной мембраны, изменяют состояние ионных каналов и скорость ферментативных реакций. НС может регулировать силу действующего раздражителя.
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
Интегрирующая роль ЦНС в организме, функции ЦНС, принципы, механизмы регуляции. Единство и особенности нервной и гуморальной регуляции функций. Нервизм (Декарт, И.М.Сеченов, С.П.Боткин, И.П.Павлов). Виды влияния нервной системы на органы и ткани.
ЦНС в организме выполняет интегрирующую роль. Она объединяет в единое целое все ткани, органы, координируя их специфическую активность в составе целостных гомеостатических и целостных функциональных систем. Интегрирующая роль осуществляется на уровне нейрона, модуля, нервного центра и взаимодействия всех отделов ЦНС, объединяющих все системы организма в единую функциональную систему.
Функции ЦНС.
Управление деятельностью опорно – двигательного аппарата. ЦНС регулирует тонус мышц и посредством его перераспределения поддерживает естественную позу, а при нарушении восстанавливает ее, инициирует все виды двигательной активности.
Регуляция работы внутренних органов. Регуляция осуществляется посредством вегетативной НС и эндокринной системы: а) в покое – обеспечение гомеостазиса; б) во время работы – приспособительная рекакция деятельности внутренних органов согласно потребностям организма и поддержание гомеостаза;
Обеспечение сознания и всех видов психической деятельности. Психическая деятельность – идеальная, субъективно осознаваемая деятельность организма, осуществляемая с помощью нейрофизиологических процессов. ВНД – совокупность нейрофизиологических процессов, обеспечивающих сознание, подсознательное усвоение инфы и целенаправленное поведение организма в окружающей среде и обществе. Низшая нервная деятельность – совокупность нейрофизиологических процессов, обеспечивающих осуществление безусловных рефлексов и инстинктов.
Формирование взаимодействия организма с окружающей средой. Осуществляется с помощью избегания или избавления от неприятных раздражителей, регуляции интенсивности обмена веществ при изменении температуры окр среды.
Регуляция функций организма – изменение интенсивности их работы для достижения полезного результата в соответствии с потребностями организма в различных условиях его жизнедеятельности. Регуляция осуществляется согласно нескольким принципам, основным из которых является системный принцип – в регуляции любого показателя организма участвует несколько органов и систем. По уровням различают клеточный, органный, системный и организменный уровни регуляции. Регуляторные механизмы могут локализоваться внутри органов и экстраорганно. Поведенческая регуляция, как и другие механизмы регуляции, направлена на поддержание гомеостаза. Имеется три механизма регуляции: нервный, гуморальный и миогенный.
Нервный механизм регуляции.
Имеется два вида влияний НС на органы – пусковое и модулирующее.
Пусковое влияние вызывает деятельность органа, находящегося в покое, прекращение импульсации, вызвавшей деятельность органа, ведет к возвращению его в исходное состояние.
Модулирующее влияние ведет к изменению интенсивности деятельности органа. Оно распространяется как на органы, деятельность которых без нервных влияний невозможна, так и на органы, которые могут работать без пускового влияния НС.
Модулирующее влияние осуществляется несколькими способами: 1. Посредством изменения характера электрических процессов в органе; 2. С помощью изменения интенсивности обмена веществ в органе; за счет изменения кровоснабжения органа.
Гуморальная и миогенная регуляция.
Гуморальная регуляция осуществляется при помощи гормонов, метаболитов и медиаторов.
Гормоны – БАВ, вырабатываемые эндокринными железами или специализированными клетками, находящимися в различных органах. Гормоны вырабатываются также нервными клетками. Это нейрогормоны, например, гормоны гипоталамуса, регулирующие функции гипофиза. БАВ вырабатываются также неспециализированными клетками – тканевые гормоны.
Тканевые гормоны – биогенные амины (гистамин, серотонин), простагландины, кинины оказывают свое действие на клетки посредством изменения их биологических свойств (проницаемости мембран, возбудимости), изменения интенсивности обменных процессов, чувствительности клеточных рецепторов, образования вторичных посредников. Они изменяют чувствительность клеток к нервным и гуморальным влияниям, поэтому их называют модуляторами регуляторных сигналов. Тканевые гормоны действуют посредством специализированных клеточных рецепторов. Тканевые гормоны изменяют проницаемость клеток, тем самым влияя на развитие ПД.
Действие гормонов или парагормонов непосредственно на соседние клетки, минуя кровь называют паракринным. Действие вещества на рецепторы клетки, выделившей это вещество, называется аутокринным.
Гормны оказывают два вида влияний: функциональное и морфогенетическое.
Функциональное влияние гормонов бывает трех видов: пусковое, модулирующее и пермиссивное. Пусковое влияние – способность гормона запускать деятельность эффектора. Модулирующее влияние – изменение интенсивности протекания процессов в органах и тканях. Модулирующим является и изменение чувствительности к действию других гормонов. Пермисивное влияние – способность одного гормона обеспечивать реализацию эффектов другого гормона.
Метаболиты – продукты, образующиеся в организме в процессе обмена веществ как результат различных биохимических реакций. Это аминокислоты, нуклеотиды, коферменты, кислоты: угольная, молочная, пировиноградная, адреналиновая, ионный сдвиг, изменение рН. Метаболиты действуют в основном как местные регуляторы, но могут влиять на другие органы и ткани, а также на активность нервных центров.
Миогенный механизм регуляции.
Сущность миогенного механизма состоит в том, что предварительное умеренное растяжение скелетной или сердечной мышц увеличивает силу их сокращения. Сократительная активность гладкой мышцы также зависит от степени наполнения полого органа, а значит, и от его растяжения.
Единство регуляторных механизмов заключается в их взаимодействии. Так, при действии холодного воздуха на терморецепторы кожи увеличивается поток афферентных импульсов в ЦНС, это ведет к выбросу гормонов, повышающих интенсивность обмена веществ, и следовательно, к увеличению теплопродукции.
Нервная и гуморальная регуляция тесно связаны, но различаются рядом свойств.
НС, в отличие от гуморального механизма регуляции, формирует ответные реакции на изменения как внешней, так и внутренней среды организма.
У нервного и гуморального механизма регуляции различные способы связи: у НС – нервный импульс как универсальный сигнал, у гуморального механизма – гормоны, медиаторы, метаболиты и тканевые гормоны.
Различается точность нервных и гуморальных механизмов. Хим. вещества попадая в кровь, разносятся по всему телу и действуют на многие органы и ткани. НС может оказывать точное локальное влияние на отдельный орган или даже группу клеток этого органа.
У нервного и гуморального механизмов различная скорость связи.
В организации гуморального механизма нередко наблюдается противоположное действие БАВ на один и тот же орган взависимости от места его воздействия
Гормональные механизмы подчиняются НС, которая передает свое влияние на эндокринные железы непосредственно или с помощью нейропептидов и своих медиаторов, выделяемых нервными окончаниями и действующих на специальные, чувствительные к медиаторам структуры – рецепторы.
Нервизм.
Почву для концепции нервизма подготовил Декарт, выдвинувший идею о рефлекторном принципе деятельности нервной системы. Гофман сформулировал гипотезу о влиянии нервов на все перемены в здоровом и больном состоянии. Согласно Куллену, все процессы в здоровом и больном организме регулирует нервный принцип, действующий через головной мозг посредством нервов – проводников нервной деятельности. По мнению Мухина, все человеческое тело можно рассматривать как построенное из нервов, ибо остальные части тела существуют вследствие нервов как управляющих их способностями.
Согласно Боткину, организм – целостная система, деятельность которой направляется и регулируется нервной системой.
Павлов обосновал представление о трофическом влиянии НС на органы и ткани, сформулировал принципы рефлекторной теории, доказал важную роль НС в регуляции секреции желез ЖКТ, ввел понятие об условных рефлексах и на их основе создал учение о ВНД.
Совокупность синаптически связанных нейронов, обеспечивающих приспособительную регуляцию функций органов или группы органов согласно потребностям организма, составляет нервный центр.
Системный принцип регуляции.
Заключается в том, что различные показатели организма поддерживаются на оптимальном уровне с помощью многих органов и систем. Системы органов объединяются в различные функциональные системы.
Функциональная система – динамическая совокупность органов и систем органов, объединяющихся для достижения организмом полезного результата.
Выделяют гомеостатические и поведенческие функциональные системы. Гомеостатические обеспечивают поддержание на оптимальных для метаболизма уровнях различных показателей организма. Это достигается посредством изменения интенсивности работы внутренних органов. Полезным результатом поведенческой функциональной системы является удовлетворение биологических потребностей, социальный и бытовой успех.
Типы регуляции функций организма.
По времени включения регуляторных механизмов относительно момента изменения величины регулируемого показателя организма имеется два типа регуляции: по отклонению и по опережению. Регуляция осуществляется с помощью обратной отрицательной связи: отклонение любого показателя от норма включает регуляторные механизмы, устраняющие это отклонение.
Регуляция по отклонению основана на циклическом механизме, при котором всякое отклонение от оптимального уровня регулируемого показателя мобилизует регуляторные механизмы для восстановления его на прежнем уровне. Регуляция по отклонению осуществляется при помощи обратной отрицательной связи, обеспечивающей разнонаправленной влияние: усиление функций органов при ослаблении показателей, и ослабление их деятельности при чрезмерном усилении и увеличения показателей организма. Положительная обратная связь оказывает только однонаправленное действие, причем стимулирует развитие процесса, находящегося под контролем управляющего комплекса.
Регуляция по опережению заключается в том, что регулирующие механизмы включаются до реального изменения показателя на основе инфы, поступающей о возможном изменении показателя в будущем.
В основе регуляции по опережению лежит механизм условного рефлекса.
Типы регуляции. Рефлекс. Универсальность и приспособленный характер изменчивости рефлекса, развитие концепции, рефлекс (Декарт, И.М.Сеченов, И.П.Павлов, П.К.Анохин).
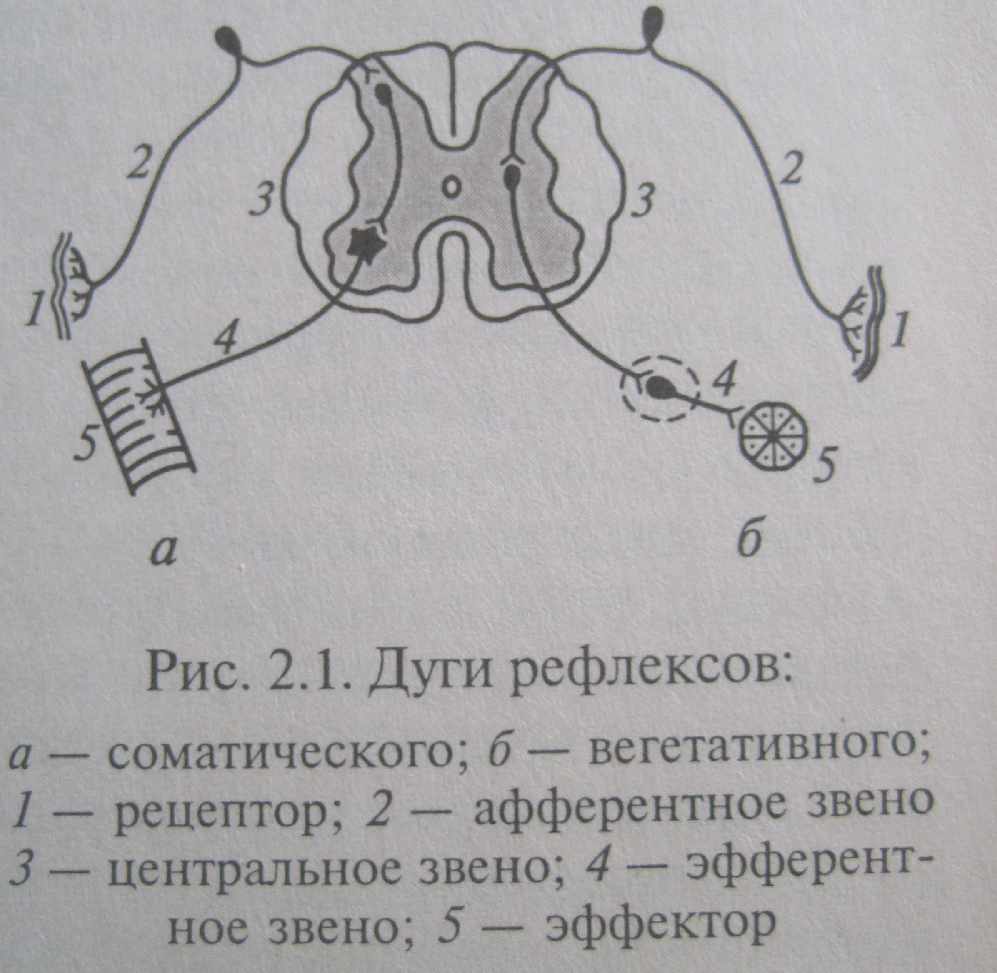
Рефлекс – ответная реакция организма на раздражение сенсорных рецепторов, осуществляемая с помощью НС. Каждый рефлекс осуществляется посредством рефлекторной дуги, которая состоит из следующих структур.
Воспринимающее звено – рецептор. Он воспринимает изменение внешней или внутренней среды, что достигается посредством трансформации энергии раздражения в рецепторный потенциал, обеспечивающий возникновение нервного импульса. Совокупность рецепторов, раздражение которых вызывает рефлекс, называется рефлексогенной зоной.
Афферентное звено, передающее сигнал в ЦНС.
Управляющее звено – совокупность центральных и периферических нейронов, формирующих ответную реакцию организма.
Эфферентное звено – аксон эффекторного нейрона. Назначение – доставка нервных импульсов к эффекторам.
Эффектор (рабочий орган).
Рефлексы классифицируют по срокам появления в онтогенезе: врожденные и приобретенные. Безусловные делят на несколько групп:
В зависимости от числа синапсов в центральной части рефлекторной дуги различают моно и полисинаптические рефлексы. Моносинаптическим является коленный разгибательный рефлекс, возникающий при ударе по сухожилию надколенника. Большинство рефлексов являются полисинаптическими, в их осуществлении участвуют несколько последовательно включенных нейронов ЦНС. От числа нейронов, участвующих в осуществлении рефлекса, зависит время рефлекса – длительность интервала от начала стимуляции до конца ответной реакции. Оно включает: время трансформации энергии раздражения в распространяющийся импульс; время проведения возбуждения в афферентном пути, в центральной части рефлекторной дуги и в эфферентном пути; время активации эффектора и его ответной реакции.
По биологическому значению: пищедобывательные, половые, защитные.
По рецепторам, раздражение которых вызывает ответную реакцию: экстероцептивные, проприоцептивные, интероцептивные.
По локализации рефлекторной дуги: центральные и периферические. Последними могут быть только вегетативные рефлексы, они подразделяются на интраорганные, межорганные и экстероорганные.
В зависимости от отдела НС: соматические и вегетативные.
Развитие концепции рефлекса.
На первом этапе были сформулированы основные положения о принципе деятельности ЦНС французским естествоиспытателем Декартом. Он сформулировал два важных положения рефлекторной теории: 1. Реакция организма на внешние воздействия является отраженной. 2. Ответная реакция на раздражение осуществляется при помощи НС.
На втором этапе было экспериментально обосновано материалистическое представление о рефлексе. Было установлено, что рефлекторная реакция может осуществляться на одном метамере лягушки. Выявлено, что стимулы могут быть не только внешними, но и внутренними; установлена роль задних чувствительных и передних двигательных корешков спинного мозга.
Сеченов обосновал приспособительный характер изменчивости рефлекса, открыл центральное торможение, а также явление суммации возбуждения в ЦНС.
На третьем этапе были открыты условные рефлексы и разработаны основы учения о ВНД. Павлов сформулировал три принципа рефлекторной теории: 1. Принцип детерминизма, согласно которому любая рефлекторная реакция причинно обусловлена – возникает при действии раздражителя; 2. Принцип структурности, суть которого заключается в том, что каждая рефлекторная реакция осуществляется при помощи определенных структур, и чем больше структурных элементов участвует в осуществлении этой реакции, тем она совершеннее; 3. Принцип единства процессов анализа и синтеза – НС анализирует с помощью рецепторов все действующие внешние и внутренние раздражители и на основании этого анализа формирует целостную ответную реакцию (синтез).
На четвертом этапе было создано учение о функциональных системах (Анохин).
Схема рефлекторной дуги соматического и вегетативного рефлексов. Общая схема функциональной системы регуляции констант организма (по П.К.Анохину).

Нейроны ЦНС: классификация, их функциональные структуры, величина потенциала покоя и потенциала действия. Классификация синапсов ЦНС, физиологическая оценка синапсов разных классов, медиаторы синапсов ЦНС и их функциональное значение.
Классификация нейронов.
По обеспечению соматических или висцеральных функций подразделяются на соматические и вегетативные соответственно двум отделам нервной системы. По месту в системе рефлекторных связей (соответственно по направлению или источнику информации) на афферентные, воспринимающие с помощью сенсорных рецепторов инфу о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС; эфферентные, передающие инфу к рабочим органам – эффекторам; вставочные, обеспечивающие взаимодействие между нейронами ЦНС. По хим природе медиатора, выделяющегося в окончаниях аксонов, адренергические, холинергические, серотонинергические и др. По типу ответной реакции эффектора тормозящие и возбуждающие. Нейроны высших отделов ЦНС по специфичности воспринимаемой сенсорной информации могут быть моно, би и полисенсорными. По характеру активности фоноактивные и молчащие, возбуждающимися только в ответ на раздражение. Фоноактивные нейроны играют большую роль в поддержании тонуса ЦНС и особенно коры большого мозга.
Классификация синапсов. В зависимости от местоположения аксо – соматические, аксодендритные, аксоаксонные, сома – соматические, дендросоматические, дендродендритные. По рабочему эффекту возбуждающие и тормозящие. По способу передачи сигналов химические, электрические, смешанные.
Свойства химических синапсов.
Большинство синапсов являются химическими. Первое их преимущество заключается в том, что хим синапсы благодаря наличию различных медиаторов и постсинаптических мембран могут оказывать возбуждающие и тормозное влияние на нервные и эффекторные клетки. Второе – могут передавать инфу в широких временных рамках. Третье – высокая пластичность; их функция зависит от предшествующей нейрональной активности, поэтому они играют важную роль в процессах обучения и памяти.
Химические синапсы. На пресинаптической поверхности активной зоны сос торны синаптической везикулы находятся филаменты и частицы, выступающие из пресинаптической терминали, часто формирующие правильный шестиугольник, названный пресинаптической решеткой. Определенные точки на сетке являются участками экзоцитоза синаптических везикул.
Медиатор в пресинаптическом окончании находится в двух пулах – готовом к экзоцитозу (пул 1 – медиатор лежит на пресинаптической мембране изнутри) и мобилизационном (пул 2 – находится в цитоскелете), из которого пополняется в течение нескольких секунд пул 1. Этоо процесс активируется ионами кальция. Химическим синапсам свойственно явление суммации.
Электросекреторное сопряжение в ЦНС обеспечивается ионами кальция: при возбуждении пресинаптической терминали открываются потенциалзависимые кальциевые каналы. Кальций входит в нервное окончание согласно электрохимическому градиенту и взаимодействует с экзоцитозным аппаратом пресинапса, представляющим собой совокупность белков.
Активация белков кальцием вызывает выброс медиатора в синаптическую щель. Синаптическая щель хим синапсов содержит поперечно расположенные гликопротеиновые филаменты, обеспечивающие связи пре и постсинаптической мембран и направление диффундирующего медиатора. Медиатор взаимодействует с белком – рецептором постсинаптической мембраны, и эффекторная клетка приходит в состояние возбуждения или торможения.
Электрические возбуждающие синапсы. Обнаружены в составе мезенцефального ядра тройничного нерва, вестибулярного ядра Дейтерса, ядра нижней оливы продолговатого мозга. Характерны для сердца, гладких мышц, секреторных клеток. Имеются следующие электрические синапсы: аксон – сома, аксон – дендрит, аксон – аксон, дендрит – дендрит, дендрит – сома, сома – сома.
Проводят сигнал в обе стороны без синаптической задержки, передача не блокируется при удалении кальция. Контактирующие мембраны клеток в составе электрического синапса связаны друг с другом полуканалами белковой природы – коннексонами. Через коннексоны клетки обмениваются аминокислотами, пептидами, РНК, метаболитами, циклическими нуклеотидами.
Механизм передачи возбуждения в электрическом синапсе: ПД, возникающий на мембране одной клетки, с помощью своего электрического поля деполяризует мембрану соседней клетки до КП.
Механизм возбуждения нейронов ЦНС: реакция нейрона на одиночный импульс и серию импульсов, возбуждающий постсинаптический потенциал (ВПСП) и его свойства, роль кальция, место возникновения ВПСП и потенциала действия, роль дендритов.
Механизм возбуждения нейронов.
В возникновении ПД нейронов принимают участие ионы кальция, ток которых в клетку более медленный, чем ток натрия (натрий вносит основной вклад в возникновение ПД).
Для возникновения ПД в нейроне необходим поток афферентных импульсов. Один ПД, пришедший в пресинаптическое окончание, обеспечивает выделение 1 – 20 квантов медиатора. При этом возникает небольшой ВПСП. Выброс медиатора обеспечивает входящий в дерполяризационную терминаль кальций. При поступлении импульсов к постсинаптическому нейрону в результате суммации ВПСП возникает ПД постсинаптического нейрона. ВПСП возникает вследствие суммарного тока ионов в клетку и из клетки через ионные каналы по электрохим градиенту.
Поступившей в пресинаптическое окончание кальций удаляется из него при помощи кальциевого насоса. Прекращение действия медиатора осуществляется посредством обратного захвата пресинаптическим аппаратом, с помощью разрушения специальными ферментами. Норадреналин разрушается монаминоксидазой и катехолметилтрансферазой, АХ гидролизуется ацетилхолинтрансферазой. Прекращение действия избытка медиатора на постсинаптическую мембрану предотвращает десенситизацию – снижение чувствительности постсинаптичекой мембраны к действующему медиатору.
Пептидные медиаторы, ферменты и другие белки транспортируются в пресинаптические окончания из тела клетки по аксону с помощью микротрубочек и микрофиламентов, тянущихся по всей длине аксона. Для этого транспорта необходимы кальций и энергия.
Место возникновения генераторных ВПСП - тело нейрона. Поскольку постсинаптические мембраны синапсов находятся в непосредственной близости от места возникновения ПД, располагающегося на аксонном холмике. Близость к аксонному холмику ВПСП, возникающих в теле нейрона, обеспечивает участие их в генерации ПД.
Место возникновения ПД нейрона: формирование ПД начинается на мембране аксонного холмика. Синапсы на нем отсутствуют, возбудимость мембраны в 3 – 4 раза выше возбудимости мембраны тела нейрона, что объясняется более высокой концентрацией натриевых каналов. ВПСП, возникающие в любых участках тела нейрона суммируются, их суммарное электрическое поле может деполяризовать мембрану на расстоянии 1 – 2 мм. Электрическое поле начального возбуждения аксонного холмика обеспечивает деполяризацию тела нейрона до КП в результате чего возникает ПД нейрона, который проводится по аксону к др клетке. На восходящей части ПД нейрона регистрируется перегиб на уровне 30 - 40 мВ (задержка в развитии ПД) вследствие того, что возбудимость мембраны тела нейрона в 3 – 4 раза ниже возбудимости мембраны аксонного холмика. ПД нейрона – единый одновременный процесс возбуждения аксонного холмика и тела нейрона.
Роль дендритов в возникновении возбуждения нейрона: дендритные синапсы называются модуляторными. Они удалены на значительное расстояние от генераторного пункта нейрона – аксонного холмика. Суммарное электрическое поле их ВПСП не может вызвать достаточную деполяризацию аксонного холмика и обеспечить возникновение ПД, а определяют лишь возбудимость нейрона.
Особенности распространения возбуждения в ЦНС, их причины, экспериментальные доказательства, клинические аспекты.
Одностороннее распространение возбуждения в нейронных цепях, рефлекторных дугах от аксона одного нейрона к телу или дендритам другого объясняется свойствами химических синапсов, которые проводят возбуждение только в одном направлении, т.к. пресинаптическая мембрана чувствительна только к электрическому импульсу, а постсинаптическая к медиатору.
Замедленное распространение возбуждения в ЦНС по сравнению с нервным волокном объясняется наличием на путях распространения возбуждения многих химических синапсов, в каждом из которых имеется синаптическая задержка около 0,5 мс. Суммарная задержка передачи возбуждения достигает 2 мс.
Дивергенция возбуждения в ЦНС объясняется ветвлением аксонов нейронов, их способностью устанавливать многочисленные связи с другими нейронами, наличием вставочных нейронов, аксоны которых также ветвятся. Дивергенцию возбуждения можно наблюдать в опыте на спинальной лягушке, когда слабое раздражение вызывает сгибание одной конечности, а сильное - энергичное движение всех конечностей или даже туловища.
Конвергенция возбуждения (принцип общего конечного пути) – схождение возбуждения различного происхождения по нескольким путям к одному и тому же нейрону или нейронному пулу (принцип воронки Шеррингтона). Объясняется наличием многих аксонных коллатералей, вставочных нейронов, а также тем, что афферентных путей в несколько раз больше, чем эфферентных нейронов.
Циркуляция возбуждения по замкнутым нейронным цепям может продолжаться несколькими минутами или даже часами. Циркуляция возбуждения – одна из причин явления последействия.
Пресинаптическое усиление: в аксо – аксональных возбуждающих синапсах возбуждение одного нервного окончания обеспечивает деполяризацию другого нервного окончания. Вследствие этого в деполяризированное окончание входит кальций, который способствует выделению медиатора
Распространение возбуждения в ЦНС легко блокируется фармакологическими препаратами, что находит широкое применение в клинической практике. В физиологических условиях ограничения распространения возбуждения по ЦНС связаны с включением нейрофизиологических механизмов торможения нейронов.
Свойства нервных центров, экспериментальные доказательства, клинические аспекты. Методы исследования ЦНС.
Свойства нервных центров.
Явление суммации возбуждения в ЦНС открыл Сеченов в опыте на лягушке: раздражение конечности слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами вызывает ответную реакцию – лягушка совершает прыжок. Значение суммации заключается в том, что нервные центры отвечают лишь на существенные для организма раздражители. Различают временную и пространственную суммацию.
Последействие - продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным путям, главной причиной которого является циркуляция возбуждения по замкнутым нейронным цепям.
Фоновая активность нервных центров объясняется: 1) спонтанной активностью нейронов ЦНС; 2) гуморальными влияниями БАВ; 3) афферентной импульсацией от различных рефлексогенных зон; 4) суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах; 5) циркуляцией возбуждения в ЦНС. Значение фоновой активности заключается в обеспечении некоторого исходного уровня деятельного состояния центра и эффекторов. Этот уровень может повышаться или понижаться в зависимости от колебаний суммарной активности нейронов нервного центра регулятора. Пейсмекерный нейрон порождает градуальные потенциалы, которые, достигая порога, запускают генерацию ПД. У такого нейрона ритмический ПД регистрируется даже при полной изоляции нейрона. Ритмическая активность мозга скорее всего представляет именно этот вид потенциалов.
Трансформация ритма возбуждения – изменение числа импульсов, поступающих к центру, относительно числа импульсов, выходящих из данного центра. Увеличению числа импульсов, возникающих в центре в ответ на афферентную импульсацию способствует дивергенция процессов возбуждения и последействие. Уменьшение объясняется снижением возбудимости нервного центра за счет процессов пре и постсинаптического торможения, а также избыточным потоком афферентных импульсов.
Высокая чувствительность ЦНС к изменениям внутренней среды. В первую очередь реагируют синапсы нейронов. При снижении содержания глюкозы в два раза могут возникать судороги.
Пластичность нервных центров связана со способностью нервных элементов к перестройке функциональных свойств. Основные проявления следующие.
Синаптическая потенциация – улучшение проведения в синапсах после кратковременной их активации, которая ведет к увеличению постсинаптических потенциалов. Может иметь пре и постсинаптическую локализацию, может быть кратковременной и долговременной.
Кратковременная после одиночных стимулов выражена слабо, после раздражающей серии потенциация в ЦНС длится дольше. Одной из причин возникновения является накопление кальция в пресинаптических окончаниях. Соответственно количество медиатора, выделяемого при каждом стимуле, увеличивается, возрастает ВПСП.
Долговременная потенциация – быстро развивающееся устойчивое повышение возбудимости постсинаптического нейрона при высокочастотных повторных или кратковременных сильных активациях пресинаптического нейрона. Связана с внутриклеточным повышением концентрации ионов кальция, вызванной активацией постсинаптических nmda – рецепторов глутаматом. Эти рецепторы являются кальциевыми каналами, пропускающими в клетку большие количества ионов кальция и натрия. В неработающем организме эти рецепторы закрыты магнием. Сильное возбуждение выбивает магниевую пробку. Вошедшие в клетку натрий и кальций вызывают значительную деполяризацию клетки, приближающуюся к КП. В этих условиях достаточно даже миниатюрных ВПСП, чтобы возникло возбуждение постсинаптического нейрона.
Значение синаптической потенциации заключается в том, что она создает предпосылки улучшения процессов переработки информации в нейронах нервных центров. Может вызвать переход центра в доминантное состояние.
Доминанта – стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе функции других нервных центров. Открыл Ухтомский в опытах с раздражением двигательных зон коры большого мозга и наблюдением сгибания конечности животного. Если раздражать корковую двигательную зону на фоне сильного повышения возбудимости другого нервного центра, обычного сгибания конечности не происходит.
Доминантный очаг возбуждения обладает следующими свойствами: стойкость, повышенная возбудимость, способность притягивать к себе иррадиирущие по ЦНС возбуждения и оказывать угнетающее воздействие на центры конкуренты и др нервные центры.
Значение доминантного очага возбуждения состоит в том, что на его базе формируется конкретная приспособительная деятельность, ориентированная на достижение полезных результатов, необходимых для устранения причин, поддерживающих тот или иной нервный центр в доминантном состоянии.
Синаптическая депрессия (утомляемость нервных центров) – ухудшение проводимости вплоть до полной блокады проведения возбуждения при длительном функционировании синапсов. Объясняется расходованием медиатора, накоплением метаболитов, закислением среды при длительном проведении возбуждения по одним и тем же нейронным цепям.
Компенсация нарушенных функций после повреждения того или иного центра – результат проявления пластичности ЦНС. Нарушенная функция частично, а иногда и полностью восстанавливается за счет большей активности сохранившихся нейронов и вовлечения в эту функцию других рассеянных нейронов в коре большого мозга с подобными функциями.
Адаптация к протезам является проявлением пластичности.
Существуют следующие методы исследования функций ЦНС:
1. Метод перерезок ствола мозга на различных уровнях. Например, между продолговатым и спинным мозгом.
2. Метод экстирпации (удаления) или разрушения участков мозга.
3. Метод раздражения различных отделов и центров мозга.
4. Анатомо-клинический метод. Клинические наблюдения за изменениями функций ЦНС при поражении ее каких-либо отделов с последующим патологоанатомическим исследованием.
5. Электрофизиологические методы:
а. электроэнцефалография – регистрация биопотенциалов мозга с поверхности кожи черепа. Методика разработана и внедрена в клинику Г. Бергером.
б. регистрация биопотенциалов различных нервных центров; используется вместе со стереотаксической техникой, при которой электроды с помощью микроманипуляторов вводят в строго определенное ядро.
в. метод вызванных потенциалов, регистрация электрической активности участков мозга при электрическом раздражении периферических рецепторов или других участков;
6. метод внутримозгового введения веществ с помощью микроинофореза;
7. хронорефлексометрия – определение времени рефлексов.
Процессы торможения в ЦНС: экспериментальные данные (И.М.Сеченов, Гольц, Мегун), механизм постсинаптического и пресинаптического торможений, их разновидности (показать с помощью схемы), значение торможения.
Торможение – активный нервные процесс , результатом которого является ослабление или прекращение возбуждения.
Торможение в ЦНС открыл Сеченов. В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Латентное время значительно увеличивается если на зрительный бугор предварительно положить кристаллик поваренной соли. Гольц обнаружил проявления торможения у спинальной лягушки. Механическое раздражение кончиков пальцев одной лапки существенно удлиняет латентное время сгибательного рефлекса другой при погружении ее в раствор кислоты. Мегун доказал наличие специальных тормозных структур в продолговатом мозге. В опытах на кошках при изучении разгибательного рефлекса он обнаружил, что раздражение медиальной части РФ продолговатого мозга тормозит рефлекторную деятельность спинного мозга.
Особенности и механизм постсинаптического торможения.
Постсинаптическое торможение обнаружил и описал Дж. Экклс при регистрауции потенциалов мотонейронов спинного мозга у кошки во время сокращения и расслабления мышцы в ходе реализации соответствующих рефлекторных актов. При рефлекторном расслаблении мышц на мотонейронах регистрируется гиперполяризационный постсинаптический потенциал, уменьшающий возбудимость мотонейрона и угнетающий его способность реагировать на возбуждающие влияния. Это ТПСП.
Амплитуда ТПСП 1 – 5 мВ. Он способен суммироваться. Тормозные синапсы постсинаптического торможения локализуются в основном на теле нейрона.
Механизм постсинаптического торможения.
Возбудимость клетки во время ТПСП понижается, поскольку увеличивается пороговый потенциал, т.к. КП остается на прежнем уровне, а мембранный потенциал возрастает. ТПСП возникает под действием аминокислоты глицина и ГАМК. В спинном мозге глицин выделяется в синапсах особых тормозных клеток (клетки Реншоу), образуемых этими клетками на мембране постсинаптического нейрона. Действуя на ионотропный рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для хлора, при этом хлор поступает в клетку согласно концентрационному градиенту, в результате развивается гиперполяризация. При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа хлора и выхода калия из клетки. Активация ГАМК1 рецепторов ведет к непосредственному увеличению проницаемости мембраны для хлора. Активация ГАМК2 рецепторов реализуется с помощью вторых посредников (цАМФ), при этом повышается проницаемость для калия.
ТПСП могут возникать вследствие уменьшения проницаемости мембраны для натрия, что сопровождается гиперполяризацией клеточной мембраны.
Разновидности постсинаптического торможения.
Выделяют возвратное, латеральное, параллельное и прямое (реципрокное) постсинаптическое торможение.
Возвратное постсинаптическое торможение – такое торможение, когда тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют. Типичным примером является торможение в мотонейронах спинного мозга.
Параллельное торможение возбуждение блокирует само себя, за счет дивергенции по коллатерали с включением тормозной клетки на своем пути и возвратом импульсов к нейрону, который активировался этим же возбуждением.
Латеральное тормозные вставочные нейроны соединены таким образом, что они активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями, в результате в соседних клетках развивается глубокое торможение.
Прямое торможение вызывает угнетение центра антагониста. Возбуждающие импульсы поступают в клетку мышцы сгибателя, а через тормозную клетку к центру мышцы антагониста.
Пресинаптическое торможение.
Было выявлено в спинном мозге в опыте с регистрацией активности мотонейронов моносинаптической рефлекторной дуги при раздражении антагонистических мышечных нервов. Раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных альфа мотонейронов. Однако опережающее раздражение афферентов сухожильных рецепторов мышц антагонистов предотвращало возбуждение активируемых альфа мотонейронов. Не изменялись мембранный потенциал и возбудимость исследуемых мотонейронов, либо регистрировался низкоамплитудный ВПСП, не способный вызвать ПД.
Механизм пресинаптического торможения.
Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях медиатором является ГАМК, который действует на ГАМК1 рецепторы.
Под действием ГАМК тормозных нейронов и последующего повышения проницаемости мембраны для хлора ионы хлора начинают выходить согласно электрическому градиенту. Это приводит к деполяризации пресинаптических терминалей и ухудшению их способности проводить импульсы.
Разновидности пресинаптического торможения.
Можно разделить на две группы: в первой блокируется собственный путь самим распространяющимся возбуждением с помощью вставочных тормозных клеток (параллельное и возвратное торможение); во второй блокируются другие нервные элементы под влиянием импульсов от соседних возбуждающих нейронов с включением тормозных клеток (латеральное и прямое).
Роль различных видов торможения.
Торможение выполняет охранительную ролью. Отсутствие торможения привело бы к истощению медиаторов в аксонах нейронов и прекращению деятельности ЦНС. Большое значение имеет для обработки поступающей в ЦНС информации. Торможение параллельных путей способствует выдеоению существенных сигналов из фона. Является важным фактором координационной деятельности ЦНС.


Координационная деятельность ЦНС: понятие о координации, что лежит в ее основе, факторы координации, примеры, их иллюстрирующие. Понятие о сегментарном и надсегментарном принципе деятельности нервных центров.
Координационная деятельность ЦНС.
Это согласование деятельности различных отделов ЦНС с помощью упорядочения распространения возбуждения между ними. Основой является взаимодействие процессов возбуждения и торможения.
Координационная деятельность обеспечивается несколькими факторами.
Фактор структурно - функциональной связи – наличие между отделами ЦНС, между ЦНС и различными органами функциональной связи, обеспечивающей преимущественной распространение возбуждения между ними.
Прямая связь – управление другим центром или рабочим органом с помощью посылки к ним эфферентных импульсов.
Обратная связь (обратная афферентация) – управление нервным центром или рабочим органом с помощью афферентных импульсов, поступающих от них самих.
Возвратная связь обеспечивает торможение нейронов вслед за их возбуждением.
Реципрокная (сочетанная) связь обеспечивает торможение центра антагониста при возбуждении центра агониста.
Принцип модульной (ансамблевой) структурно – функциональной организации ЦНС: каждый модуль представляет собой совокупность повторяющихся локальных нейронных связей, обрабатывающих и передающих инфу с помощью внутренних и внешних связей. Основным функциональным признаком модульной организации является локальный синергизм реакций нейронов центральной части ансамбля, окруженной зоной заторможенных нейронов – тормозная оконтовка.
Фактории субординации – подчинение нижележащих отделов ЦНС вышележащим.
Фактор силы. В случае одномоментной активации путей от различных рефлексогенных зон, центр реагирует на более сильный.
Одностороннее проведение возбуждения в химических синапсах ЦНС способствует упорядоченному распространению возбуждения, ограничивая иррадиацию возбуждения в ЦНС.
Синаптическая потенциация участвует в процессах координационной деятельности в процессах выработки навыков.
Доминанта. Доминантное состояние двигательных центров обеспечивает автоматизированное выполнение двигательных актов.
Роль спинного мозга в регуляции функций организма: вегетативные и соматические центры и их значение. Состояние тонуса мышц спинального животного. Спинальный шок. Значение афферентной и эфферентной импульсации.
Соматические нейроны спинного мозга. Нейроны спинного мозга классифицируют по принадлежности к отделам НС – соматические и вегетативные; по назначению – эфферентные, афферентные, вставочные, ассоциативные; 3)по влиянию – возбуждающие и тормозные.
Эфферентные нейроны относятся к соматической нс, являются эффекторными, поскольку иннервируют непосредственно рабочие органы – эффекторы (скелетные мышцы), их называют мотонейроны. Различают альфа и гама мотонейроны. Альфа мотонейроны иннервируют экстрафузальные мышечные волокна (скелетная мускулатура), их аксоны характеризуются высокой скоростью проведения возбуждения. Они подразделяются на две подгруппы альфа1 мотонейроны – быстрые, иннервирующие белые мышечные волокна, и альфа2 – медленные, иннервирующие красные мышечные волокна.
Гамма – мотонейроны рассеяны среди альфа мотонейронов, их активность регулируется нейронами вышележащих отделов ЦНС. Они обладают спонтанной активностью, иннервируют интрафузальные мышечные волокна мышечного веретена (мышечного рецептора).
Афферентные нейроны локализуются в спинальных ганглиях и ганглиях черепных нервов. Их отростки, проводящие афферентную импульсацию от мышечных, сухожильных и кожных рецепторов, вступают в ствол мозга или соответствующие сегменты спинного мозга и образуют синаптические контакты либо на альфа мотонейронах, либо на вставочных нейронах.
Вставочные нейроны устанавливают связь с мотонейронами спинного мозга чувствительных нейронов. Они обеспечивают связь спинного мозга с ядрами ствола , а через них с корой большого мозга.
Ассоциативные нейроны образуют собственный аппарат спинного мозга, устанавливающий связь между сегментами и внутри одного сегмента. Участвует в поддержании позы, тонуса мышц, движений.
Нейроны РФ. РФ спинного мозга состоит из тонких перекладин серого вещества, пересекающихся в различных направлениях, ее нейроны имею большое количество отростков. РФ обнаруживается на уровне шейных сегментов между передними и задними рогами, а на уровне верхнегрудных сегментов – между боковыми и задними рогами в белом веществе, примыкающем к серому.
Спинальные нейроны вегетативной нервной системы. Неуроны симпатической нервной системы являются вставочными. Расположены в боковых рогах грудного, поясничного и частично шейного отделов. Фоноактивны. Нейроны парасимпатической НС также вставочные и фоноактивные, локализуются в сакральном отделе спинного мозга (S2 – S4).
Центры спинного мозга. Различные центры симпатического отдела НС локалихзованы в: центр зрачкового рефлекса – C8 – Th12; центр регуляции деятельности сердца – Th1 – Th5; слюноотделения – Th2 – Th4; регуляции функции почек – Th5 – L3; сегментарно расположены центры регулирующие функции половых желез и сосудов, гладких мышц внутренних органов, центры пиломоторных рефлексов. Парасимпатическую иннервацию (S2 – S4) получают все органы малого таза: мочевой пузырь, часть толстой кишки ниже ее левого изгиба, половые органы.
Центры управления скелетной мускулатурой находятся во всех отделах спинного мозга и иннервируют по сегментарному принципу скелетную мускулатуру шеи, диафрагмы, верхних конечностей, туловища и нижних конечностей.
Проводниковая функция спинного мозга осуществляется нисходящих и восходящих путей.
Афферентная инфа поступает в спинной мозг в основном через задние корешки, эфферентная импульсация в передних корешках и регуляция функций различных органов и тканей организма осуществляется через передние корешки (закон Белла – Мажанди).
Все афферентные входы в спинной мозг несут инфу от трех групп рецепторов: 1) кожных – болевых, температурных, прикосновения, давления, щекотки, вибрации; 2) проприорецепторов мышц (мышечных веретен), сухожилий (рецепторов Гольджи), надкостницы и оболочек суставов; 3) от рецепторов внутренних органов – висцерорецепторов (механо и хеморецепторов).
Медиатором первичных афферентных нейронов является глутамат, модулятором – субстанция П, энкефалин, ВИП.
Значение афферентной импульсации заключается в следующем: 1) участвует в координационной деятельности ЦНС по управлению скелетной мускулатурой; при выключении афферентной импульсации от рабочего орган управление им становится несовершенным; 2) участвует в процессах регуляции функций внутренних органов; 3) поддерживает тонус ЦНС, при выключении афферентной импульсации происходит уменьшение суммарной тонической активности ЦНС; 4) афферентная импульсация несет в вышележащине отделы ЦНС инфу об изменениях окружающей среды.
Тонус мышц у спинального организма.
Тонус формируют статические тонические рефлексы мышц сгибателей и разгибателей.
Сразу после перерезки или при повреждении спинного мозга у человека наблюдается мышечная атония и отсутствие рефлексов (спинальный шок).
Главной причиной является выключений влияния на спинной мозг вышележащих отделов ЦНС. Фазные сгибательные и разгибательные рефлексы нижних конечностей у человека в случае повреждения спинного мозга усиливаются.
Гипертонус имеет рефлекторную природу, он развивается вследствие афферентной импульсации от мышечных рецепторов.
Имеется два фактора, обеспечивающих афферентную импульсацию от мышечных рецепторов по задним корешкам спинного мозга, которая ведет к возбуждению альфа мотонейронов спинного мозга и сокращению скелетных мышц. 1) спонтанная активность мышечных рецепторов, поступление афферентных импульсов к альфа мотонейронам, последние возбуждаются и посылают импульсы к мышцам, повышая тонус. 2) спонтанная активность гамма мотонейронов. Возбуждение гамма мотонейронов вызывает возбуждение и сокращение иннервируемых ими интарфузальных мышечных волокон, в результате чего увеличивается натяжение мышечного рецептора, поскольку концы его прикреплены к скелетной мышце. Вследствие натяжения мышечного рецептора раздражаются и возбуждаются афферентные окончания (проприорецепторы), импульсы от которых поступают к альфа мотонейронам и возбуждают их. Альфа мотонейроны посылают импульсы к скелетной мышце, вызывая ее постоянное (тоническое) сокращение.
Соматические рефлексы спинного мозга: их характеристика, классификация по характеру ответной реакции (рефлексогенные зоны и функциональное значение каждого из рефлексов), механизм шагательного рефлекса (схема).
Классификация соматических рефлексов спинного мозга.
По иду рецепторов, раздражение которых вызывает рефлекс: проприорецептивные, висцерорецептивные, кожные. Последниф являются защитными. Рефлексы, возникающие с проприроепторов, участвуют в акте ходьбы и регуляции мышечного тонуса, что важно для поддержания позы организма. Висцеререцептивеные рефлексы возникают с интерорецепторов и проявляются в сокращении мышц брюшной стенки, грудной клетки и разгибателей спины.
По органам (эффекторам рефлекса): рефлексы конечностей, брюшные, области таза.
Рефлексы конечностей могут быть фазными и тоническими.
Фазные рефлексы подразделяются на сгибательные, разгибательные и ритмические.
Сгибательные рефлексы конечностей – однократное сгибание конечности при однократном раздражении кожи или проприорецепторов. Одновременно с возбуждением мотонейронов мышц сгибателей происходит торможение мотонейронов мышц разгибателей. Кожные рефлексы являются полисинаптическими, имеют защитный характер. Проприорецептивные моносинаптические. Они участвуют в формировании акта ходьбы.
Разгибательные рефлексы возникают только с проприорецепторов мышц разгибателей и являются моносинаптическими. Мотонейроны мышц сгибателей во время разгибательного рефлекса тормозятся – постсинаптическое реципрокное торможение с участием вставочных клеток.
Ритмические рефлексы конечностей – многократное повторное сгибание и разгибание конечностей.
Тонические рефлексы конечностей включают две группы: сегментарные и надсегментарные.
Сегментарные тонические рефлексы. Главное назначение в поддержании позы. Тоническое сокращение скелетных мышц является фоновым для осуществления всех двигательных актов. В положении стоя тоническое сокращение мышц разгибателей предотвращает сгибание нижних конечностей и обеспечивает сохранение вертикальной естественной позы. Тонические рефлексы на растяжение мышц называются миотатическими.
Надсегментарные тонические рефлексы. Возникают при наклоне головы вверх или вниз, а также при поворотах и наклонах головы влево и вправо. при наклоне головы вниз увеличивается тонус мышц сгибателей передних конечностей и тонус мышц разгибателей задних конечностей, в результате передние конечности сгибаются, а задние разгибаются. При наклоне головы вверх возникают противоположные реакции – передние конечности разгибаются вследствие увеличения тонуса их мышц разгибателей, задние сгибаются вследствие повышения тонуса их мышц сгибателей. Эти рефлексы возникают с проприорецепторов мышц шеи и фасция, покрывающих шейный отдел позвоночника. При поворотах головы влево или вправо повышается тонус мышц разгибателей обеих конечностей на стороне, куда повернута голова, и повышается тонус мышц сгибателей на противоположной стороне. Рефлекс направлен на сохранение позы, которая может быть нарушена вследствие смещения центра тяжести в сторону поворота головы.
Брюшные рефлексы. Верхний, нижний, средний брюшные рефлекс являются фазными. Они выражаются в сокращениях соответствующих участков мускулатуры стенки живота. Это защитные рефлексы.
Рефлексы области таза. К ним относятся кремастерный (яичковый) и анальный рефлексы. Оба фазные.
Шагательный рефлекс.
Рефлекс вызывают однократным раздражением кожи конечности. Он выражается в сгибании этой конечности с одновременным разгибанием противоположной задней конечности – перекрестный разгибательный рефлекс. Затем согнутая конечность разгибается, опускается вниз, а разогнутая сгибается и поднимается вверх и т.д.
Значение проприорецепторов в шагательном рефлексе.
Мышечные веретена (мышечные проприорецепторы) расположены параллельно скелетной мышце: своими концами крепятся к соединительно тканной оболочке пучка экстрафузальных мышечных волокон при помощи напоминающих сухожилия волокон соединительной ткани. Поэтому, когда мышца расслабляется (удлиняется), растягиваются и мышечные рецепторы, что и ведет к их возбуждению.
Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон, окруженных соединительнотканной капсулой.
Вокруг средней части мышечного веретена обвивается несколько раз окончание одного афферентного волокна. Многие мышечные веретена иннервируются и другим афферентным волокном, которое также обвивает в виде спирали мышечный рецептор, но периферические его участки. Импульсы от мышечных рецепторов возбуждают нейроны своего двигательного центра и тормозят нейроны центра антагониста. Ближе к концам мышечных веретен подходят двигательные нервные окончания, аксоны гамма мотонейронов спинного мозга. Их импульсация вызывает сокращение интарфузальных мышечных волокон, что ведет к возбуждению рецептора.
Сухожильные рецепторы (рецепторы Гольджи) заключены в соединительнотканную капсулу и локализуются в сухожилиях скелетных мышц вблизи от сухожильно – мышечного соединения. Они представляют собой безмиелиновыее окончания толстого миелинового афферентного волокна. Крепятся относительно скелетной мышцы последовательно, что обеспечивает их раздражение и возбуждение при натяжении сухожилий. Сухожильные рецепторы посылают в мозг инфу о том, что мышца сокращена, тогда как мышечные рецепторы сигнализируют о том, что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра антагониста.
Механизм шагательного рефлекса.
Поочередное сокращение и расслабление скелетных мышц запрограммированы в спинном мозге, важную роль играет импульсация от проприорецепторов в двигательный центр к каждой мышце.
Когда мышца расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от них поступают к своим альфа мотонейронам спинного мозга и возбуждают их. Далее альфа мотонейроны посылают импульсы к той же мышце, что ведет к ее сокращению. Как только мышца сократилась, возбуждение мышечных веретен прекращается или сильно уменьшается и начинают возбуждаться сухожильные рецепторы. Импульсы от последних поступают в свой спинальный центр, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение альфа мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление снова возбуждает мышечные рецепторы и альфа мотонейроны и мышца вновь сокращается.
Шагательные координированные движения возможны у спинального животного в отсутствие обратной афферентации проприорецепторов, благодаря наличию генератора локомоторного цикла (каждая конечность может совершать самостоятельные движения) при этом важную роль играют межсегментарные связи на уровне спинного мозга.

Продолговатый мозг и мост: центры и соответствующие им рефлексы, их отличия от рефлексов спинного мозга, состояние мышечного тонуса бульбарного животного, схема, объясняющая механизм его происхождения.
Непосредственным продолжением вверх спинного мозга у всех позвоночных животных и человека является продолговатый мозг. Вместе с варолиевым мостом продолговатый мозг является производным ромбовидного мозга (rhombencephalon). Одновременно он является частью ствола мозга (truncus encephalicus). Ствол - это осевые структуры головного мозга, включающие продолговатый мозг, мост, средний и промежуточный мозг.
В продолговатом мозге и мосту по сравнению со спинным мозгом нет четкой сегментации серого и белого вещества. Скопления нейронов здесь формируют ядра, обеспечивающие сложную рефлекторную деятельность.
В продолговатом мозге и варолиевом мосту (в дорзальной их части) находится большая группа ядер черепных нервов (с V по XII пары), которые образуют массу серого вещества ромбовидной ямки. Эти ядра являются жизненно важными функциональными центрами. На дне IV желудочка в продолговатом мозге находится дыхательный центр, состоящий из центра вдоха и центра выдоха. Нейроны этих центров посылают импульсы к дыхательным мышцам через мотонейроны спинного мозга, контролируя правильное чередование фаз дыхательного цикла. Рядом лежит сосудодвигательный центр (контролирует тонус стенок сосудов и уровень АД) и сердечно-сосудистый центр (согласует состояние сосудов с работой сердца).
В продолговатом мозге также лежат пищеварительные центры:
1 - моторные (жевания, глотания, моторики желудка и части кишечника),
2 - секреторные (слюноотделения, желудочной секреции, выделения соков тонкой кишки, поджелудочной железы и др.).
Здесь же находятся центры защитных рефлексов (чихания, кашля, рвоты, мигания, слезоотделения). Таким образом, биологическая роль продолговатого мозга заключается в регуляции постоянства состава внутренней среды организма (гомеостаза). Он также осуществляет более тонкие приспособительные реакции организма к внешней среде, чем спинной мозг.
Кроме ядер черепных нервов в продолговатом мозге имеются переключательные чувствительные ядра. В толще олив лежат крупные нижние оливные ядра, а также медиальные и верхние добавочные оливные ядра. На задней поверхности в одноименных бугорках залегают тонкое ядро Голля и клиновидное ядро Бурдаха.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса мышц. Импульсы от вестибулярных ядер (Дейтерса и Бехтерева) усиливают тонус мышц разгибателей, что необходимо для организации позы. Неспецифические отделы продолговатого мозга (ядра РФ) наоборот снижают тонус мышц, в том числе и разгибателей. Продолговатый мозг участвует в осуществлении рефлексов поддержания и восстановления позы тела (установочных рефлексов).
Продолговатый мозг выполняет две функции (рефлекторную и проводниковую). Рефлекторная функция осуществляется за счет:
1 - простых сегментарных рефлексов (защитный мигательный, слезоотделения, движений ушной раковины, кашля, чихания, рвоты),
2 - лабиринтных рефлексов (распределение тонуса между отдельными группами мышц и установки определенной позы),
3 - установочных рефлексов (поддержания позы и рабочих движений),
4 - вегетативных рефлексов (дыхания, кровообращения, пищеварения).
Проводниковая функция осуществляется путем проведения:
1 - восходящих волокон от спинного мозга к коре полушарий,
2 - нисходящих волокон от коры полушарий к спинному мозгу,
3 - собственных проводящих пучков продолговатого мозга и моста, соединяющих ядро и оливу вестибулярного нерва с мотонейронами спинного мозга.
Продолговатый мозг принимает чувствительные волокна от рецепторов мимических и жевательных мышц, мышц шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, полости носа и рта, от рецепторов органов слуха и равновесия, от рецепторов гортани, трахеи, легких, интерорецепторов ЖКТ и сердечно-сосудистой системы. Здесь волокна переключаются на другие нейроны, образуя путь в таламус и кору полушарий. Восходящие пути кожно-мышечной чувствительности перекрещиваются на уровне продолговатого мозга так же, как и большая часть пирамидных (двигательных) путей.
Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего мозга.
Бульбарный организм характеризуется децеребрационной ригидностью. В таком организме сохранены шейные и лабиринтные тонические рефлексы, но отсутствуют выпрямительные рефлексы и произвольные движения.

Средний мозг: основные структуры и их функции, статические и статокинетические рефлексы и опыты, их доказывающие (Магнус). Состояние мышечного тонуса мезенцефального животного, схема, объясняющая механизм его регуляции. Функции черной субстанции.
Средний мозг — отдел головного мозга, древний зрительный центр. Включен в ствол головного мозга.
Вентральную часть составляют массивные ножки мозга, основную часть которых занимают пирамидные пути. Между ножками находится межножковая ямка, из которой выходит III (глазодвигательный) нерв. В глубине межножковой ямки — заднее продырявленное вещество.
Дорсальная часть — пластинка четверохолмия, две пары холмиков, верхние и нижние. Верхние, или зрительные холмики несколько крупнее нижних (слуховых). Холмики связаны со структурами промежуточного мозга — коленчатыми телами, верхние — с латеральными, нижние — с медиальными.
Внутри нижних холмиков находятся слуховые ядра, туда идет латеральная петля. Вокруг сильвиева водопровода — центральное серое вещество.
В глубине покрышки среднего мозга (под четверохолмием) находятся ядра глазодвигательных нервов, красные ядра, чёрное вещество, ретикулярная формация.
В латеральных отделах среднего мозга в него входят верхние мозжечковые ножки, которые, постепенно погружаясь в него, образуют перекрест у средней линии. Дорсальная часть среднего мозга, расположенная кзади от водопровода, представлена крышей с ядрами нижних и верхних холмиков.
Ядра нижних холмиков играют существенную роль в реализации функции слуха и формировании сложных рефлексов в ответ на звуковые раздражения.
Ядра участвуют в осуществлении «автоматических» реакций, связанных со зрительной функцией, то есть безусловных рефлексов в ответ на зрительные раздражения. Кроме того, эти ядра координируют движения туловища, мимическую реакцию, движения глаз, головы, ушей и проч. в ответ на зрительные стимулы. Осуществляются эти рефлекторные реакции благодаря покрышечно-спиномозговому и покрышечно-бульбарному путям.
Вентральнее от верхних и нижних холмиков крыши находится водопровод среднего мозга, окружённый центральным серым веществом. В нижнем отделе покрышки среднего мозга располагается ядро блокового нерва, а на уровне среднего и верхнего отделов — комплекс ядер глазодвигательного нерва.
Важными образованиями среднего мозга являются также красные ядра и чёрная субстанция. В красных ядрах оканчиваются волокна передних мозжечковых ножек, корково-красноядерные волокна и волокна из образований стриопаллидарной системы. В красном ядре начинаются волокна красноядерно-спинномозгового, а также красноядерно-оливного путей, волокна, идущие в кору большого мозга. Таким образом, красное ядро является одним из центров, участвующих в регуляции тонуса и координации движений. В чёрном веществе берет своё начало нигростриарный путь, дофаминэргические волокна которого управляют функцией стрии.
Основание ножки среднего мозга состоит из волокон, которые соединяют кору большого мозга и другие образования конечного мозга с нижележащими образованиями мозгового ствола и спинного мозга. Большая часть основания занята волокнами пирамидного пути.
Средний мозг выполняет следующие функции:
центр ориентировочного рефлекса
центр позы
центр обработки первичной информации (зрение, слух)
регулирующую в продолжительности актов жевания и глотания
Черная субстанция ограничивает влияние норадренергической системы, активирует продукцию оксида азота в головном мозге, увеличивает устойчивость к стрессорным повреждениям. Нейроны черного вещества оказывают модулирующее влияние на фоновую активность нейронов полосатого тела. Также оказывает тормозящее действие на нейроны таламуса.
Мезенцефальный организм (нарушение связи между средним и промежуточным мозгом) имеет нормальный мышечный тонус, сохраняются все тонические рефлексы, но произвольные движения отсутствуют.
Тонические рефлексы организма Магнус разделил на две группы: статические (перераспределение мышечного тонуса при изменении положения тела в пространстве, или отдельных его частей относительно друг друга, например восстановление нарушенной позы) и статокинетические (перераспределение мышечного тонуса при перемещении организма в пространстве, например при беге, подъеме в лифте).
Статические рефлексы – тонические рефлексы организма в состоянии покоя, в том числе и при поворотах головы и движениях рук, а в положении сидя ног, вызывающих смещение центра тяжести. К ним относятся сегментарные и надсегментарные (шейные) тонические рефлексы спинного мозга и рефлексы ствола мозга, возникающие с вестибулорецепторов. Их роль – сохранение естественной позы.
Для выявления роли шейных рефлексов Магнус удалял у животного вестибулярный аппарат, чтобы погасить вестибулярные рефлексы.
Для выявления вестибулярных рефлексов на шею животного, расположенную на одной линии с туловищем, накладывают гипсовую повязку, чтобы прекратить шейные надсегментарные рефлексы. Вестибулярные рефлексы выявляются при поворачивании животного через ось висков. Если животное располагается горизонтально спиной и затылком вниз и до 45 градусов выше горизонтали, то тонус мышц разгибателей становится максимальным; если при дальнейшем повороте оно располагается спиной и затылеом вверх по горизонтали и до -45 градусов по вертикали, то тонус мышц разгибателей становится максимальным.
Статокинетические рефлексы – рефлекторное перераспределение мышечного тонуса, возникающее с кожных, проприо и вестибулорецепторов, при нарушении позы. К ним относятся выпрямительные рефлексы – выпрямление головы и туловища. Выпрямление головы запускается с вестибулярных и кожных рецепторов, выпрямление туловища с проприорецепторов шеи и кожных рецепторов.
Ретикулярная формация: расположение в ЦНС, связи ее ядер, особенности свойств нейронов, восходящие влияние ретикулярной формации и опыты, их доказывающие, типы вызванных ответов.
РФ образована нейронами, расположенными диффузно и в виде ядер в центральных отделах ствола мозга. Они имеют хорошо ветвящиеся аксоны, которые позволяют одному нейрону устанавливать связи с 25 тыс. других нейронов; принимают импульсы от сенсорных путей, идущих от разных рецепторов.
Ретикулярные нейроны полимодальные; имеют большие рецепторные поля; у них длительные латентный период ответа на периферическую стимуляцию, связанный с проведением возбуждения до них через многочисленные синапсы. Они имеют фоновую тоническую активность, высокие возбудимость и мобильность, чувствительны к некоторым веществам крови (адреналин, СО2) и лекарствам (барбитураты, аминазин)
РФ имеет афферентные входы преимущественно от трех источников: 1. От температурных и болевых рецепторов по волокнам спиноретикулярного пути тройничного нерва; 2. От сенсорной и частично других зон коры головного мозга по кортикоретикулярным путям импульсация поступает в ядра, дающие начало ретикулярным путям, а также ядрам, которые проецируются на мозжечок; 3. От ядер мозжечка по мозжечковоретикулярному пути.
Эфферентные выходы: 1. В спинной мозг – по медиальному и латеральному ретикулоспинальным путям; 2. К верхним отделам головного мозга (неспецифическим ядрам таламуса, заднему гипоталамусу, полосатому телу) идут восходящие пути, начинающиеся в ядрах продолговатого мозга и моста; 3. К мозжечку идут пути, которые начинаются в латеральном и парамедианном ретикулярном ядрах и я ядре покрышки моста.
Восходящее влияние на большой мозг. РФ может оказывать как активирующее, так и тормозное действие. Оно образуется аксонами 5 – 17 % нейронов РФ, формирующими ретикулоталамические пути. Их стимуляция приводит к появлению постсинаптических потенциалов в нейронах и вызванных потенциалов в неспецифических и специфических ядрах таламуса. После переключения в нейронах таламуса сенсорный поток проецируется в различные области коры. Восходящие влияния поступают в задний таламус, полосатое тело, прозрачную перегородку. В восходящей РФ выделяют две части: ростральную, образуемую нейронами РФ среднего мозга, и каудальную, формируемую нейронами РФ продолговатого мозга и моста.
Активирующее влияние ростральной части РФ впервые было показано с помощью перерезки ствола мозга между верхним и нижним холмиками четверохолмия. В результате перерезки животное погружалось в сноподобное состояние: нарушен контакт с внешним миром, не реагирует на световые и обонятельные раздражители. Прямое доказательство активирующего влияния РФ на большие полушарие было получено в опытах с раздражением РФ через погружные электроды у спящих животных (Мегун, Моруцци). Стимуляция РФ вызывала пробуждение животного. У него возникала ориентировочная реакция, в корковых нейронах возникали ВПСП; альфа ритм и более медленные ритм сменялись бета ритмом – реакция десинхронизации ЭЭГ.
Активирующее влияние на кору осуществляется в основном при помощи холинергических нейронов.
Тормозное влияние возникало при стимуляции каудального отдела РФ. При этом на корковых нейронах формировались в основном ТПСП и выявлялся синхронизирующий эффект на ЭЭГ. Тормозное влияние вызывает синхронизацию ЭЭГ в фазе медленного сна и атонию мышц в фазе быстрого сна. У бодрствующих животных стимуляция каудальной РФ угнетает локомоторную активность, но существенно не влияет на рефлексы пищевого поведения. Тормозные влияния осуществляются также при помощи холинергических нейронов.
Нисходящие влияния ретикулярной формации и опыты, их доказывающие. Схема, отражающая участие ретикулярной формации в регуляции мышечного тонуса.
Нисходящие влияние на тонус мышц и формирование позы РФ оказывает через латеральный и медиальный ретикулоспинальные тракты.
Медиальный ретикулоспинальный путь идет от ретикулярных ядер моста и возбуждает спинальные альфа и гамма нейроны мышц – разгибателей осевой мускулатуры (туловища, проксимальных отделов конечностей), через тормозные интернейроны тормозятся мотонейроны сгибателей.
Латеральный ретикулоспинальный путь идет от ретикулярного гигантоклеточного ядра продолговатого мозга, возбуждаются альфа и гамма нейроны мышц сгибателей мускулатуры конечностей и реципрокно затормаживают мышцы – разгибатели.
Взаимоотношения между стволовыми и двигательными центрами выявляются в эксперименте с перерезкой ствола по передней границе четверохолмия. У подопытных животных в этом случае повышается тонус мышц разгибателей конечностей, туловища, шеи, хвоста – децеребрационная ригидность.

Мозжечок: афферентные и эфферентные связи, роль мозжечка в регуляции тонуса мышц в обеспечении двигательной активности.
Выделяют три структуры мозжечка. Древний мозжечок состоит из клочка и узелка, а также часть червя. Старый мозжечок включает центральную дольку, вершину пирамиду, язычок червя, и четырехугольную дольку полушарий. Новый мозжечок состоит из полушарий и части червя (скат, листок, бугор).
Клетки Пуркинье, образующие средний слой коры, являются главной функциональной единицей с ярко выраженной интегративной направленностью. Она реализуется сильно ветвящимися дендритами, на которых в одной клетке может быть до 100 тыс синапсов. Клетки Пуркинье являются единственными эфферентными нейронами коры мозжечка, связывая ее с внутримозжечковыми и вестибулярными ядрами. К коре мозжечка проходит три вида волокон – лиановые, моховидные и моноаминергические.
Лиановые волокна являются аксонами нейронов нижней оливы продолговатого мозга. Инфа к ним поступает от мышечных, кожных рецепторов и двигательной коры головного мозга. Каждое лиановидное волокно устанавливает контакт с дендритами одной клеткой Пуркинье, но число синапсов может достигать 300, что объясняет исключительно сильное возбуждающее действие этого афферентного входа. Также они оказывают тормозящие действие на клетки Пуркинье через корзинчатые и звездчатые клетки поверхностного слоя мозжечка.
По моховидным волокнам в кору мозжечка поступает инфа от коры больших полушарий , проприорецепторов, вестибулярных рецепторов и РФ. Они образуют синапсы на дендритах клеток зерен внутреннего слоя коры. Через аксоны клеток – зерен моховидные волокна оказывают возбуждающее влияние на клетки Пуркинье и опосредованно тормозное действие через корзинчатые и звездчатые клетки. Активность клеток – зерен регулируется через тормозные клетки Гольджи внутреннего слоя по типу возвратного торможения (медиатор ГАМК).
Монаминергический афферентный вход. Через него на мозжечок передается влияние голубого пятна (медиатор норадреналин) и ядер шва (серотонин). Стимуляция этих ядер тормозит фоновую активность клеток Пуркинье и модулирует их ответы на активацию моховидных и лиановидных афферентов.
На клетки Пуркинье поступает в основном возбуждающее влияние. Но их возбуждение оказывает тормозное влияние на три парных мозжечковых ядра (ядра шатра, зубчатые и промежуточные), иннервирующих моторные центры головного мозга. Мозжечок осуществляет регуляцию по механизму сдерживания активности моторных центров.
Три парных ядра мозжечка образуют три главных эфферентных выхода на стволовые и корковые моторные центры. Нейроны этих ядер имеют высокую фоновую активность. Они находятся под тормозящим влиянием клеток Пуркинье и возбуждающим влиянием афферентных входов в мозжечок, которые по коллатеральным ответвлениям поступают в эти ядра.
Двигательные функции мозжечка связаны с координацией движений и обучением двигательным навыкам.
Регуляция мышечного тонуса, обеспечивающего поддержание естественной позы (равновесия) осуществляется преимущественно древним и частично старым мозжечком. Получая и обрабатывая импульсацию от вестибулярных рецепторов, проприорецепторов аппарата движения, кожных, зрительных и слуховых рецепторов, мозжечок способен оценить состояние мышц, положение тела в пространстве и через ядра шатра, а также прямого выхода на вестибулярные ядра изменить позу тела и сохранить равновесие. Мозжечок осуществляет тонкую настройку вестибулярных рефлексов, в том числе рефлекторное поддержание вертикальной позы.
Координация движений осуществляется старым и новым мозжечком. В кору этой части мозжечка поступает инфа от рецепторов аппарата движения, а также импульсация от моторной коры. Анализируя инфу о программе и выполнении движения, мозжечок способен через свои промежуточные ядра, имеющие выходы на красное ядро и моторную кору, осуществить координацию, а при необходимости и коррекцию движений. При этом регулируются амплитуда, направление и темп движения.
Программирование произвольных движений осуществляется корой нового мозжечка, которая получает инфу о замысле движения из ассоциативных зо коры большого мозга через ядра моста. В коре нового мозжечка она перерабатывается в программу движения, которая через зубчатое ядро мозжечка и вентральное латеральное ядро таламуса поступает в премоторную зону. Там она получает дальнейшую обработку и через пирамидную и экстрапирамидную системы реализуется как сложное целенаправленное движение. Контроль коррекция и координация медленных програмированных движений осуществляется на основе обратной афферентации от проприорецепторов, а быстрых путем изменения программы движения без сличения с обратной афферентацией.
Промежуточный мозг: структуры и их функции. Пирамидная и экстрапирамидная системы: главные структуры функции. Стриопалидарная система, и ее функции.
Промежуточный мозг расположен между средним и конечным мозгом. Он состоит из таламической области и гипоталамуса. Таламическая область охватывает ядра таламуса, включая коленчатые тела и подушки, и эпиталамус (эпифиз).
Таламус (зрительный бугор) представляет собой парный ядерный комплекс, включающий до 60 ядер, в котором выделяют три главные группы ядер: релейные (специфические, переключательные), ассоциативные и неспецифические. Все группы ядер обладают в разной степени тремя общими функциями: переключающей, интегративной и модилирующей.
Релейные ядра разделяют на сенсорные и несенсорные. Сенсорные переключают потоки афферентной импульсации в сенсорные зоны коры. В них также происходит перекодирование и обработка инфы. Выделяют три основные ядерные структуры.
Вентральные задние ядра (вентробазальный комплекс) – главное реле для переключения соматосенсорной афферентной системы, импульсы которой поступают по волокнам медиальной, спинно – мозговой и тройничной петлям. В них переключаются проприоцептивная, тактильная, вкусовая, висцеральная, частично температурная и болевая чувствительность. В этих ядрах имеется топографическая проекция периферии; при этом функционально более тонко организованные части тела имеют большую зону представительства. Импульсация из вентральных задних ядер проецируется в соматосенсорную кору постцентральной извилины, где формируются соответствующие ощущения. Электростимуляция вентральных задних ядер вызывает парестезии (ложные ощущения) в разных частях тела, иногда нарушение схемы тела.
Латеральное коленчатое тело является реле для переключения зрительной импульсации в затылочную кору, где она используется для формирования зрительных ощущений.
Медиальное коленчатое тело является реле для переключения слуховой и, возможно, вестибулярной импульсации в височную кору.
Переключательную функцию в сенсорных ядрах обеспечивают таламокортикальные (релейные) нейроны, длинный аксон которых непосредственно иннервируют нейроны сенсорной коры. Регуляция этой передачи осуществляется с помощью тормозных и возбуждающих интернейронов ядер.
Несенсорные релейные ядра таламуса (передние и вентральные) переключают в кору несенсорную импульсацию, поступающую в таламус из разных отделов головного мозга. В передние ядра импульсация в основном поступает из мамиллярных тел гипоталамуса. Нейроны передних ядер проецируются в лимбическую кору. От нее аксонные связи идут к гиппокампу и опять к гипоталамусу, в результате чего образуется нейронный круг, движение возбуждения по которому обеспечивает формирование эмоций.
Вентральные ядра участвуют в регуляции движения, выполняя таким образом моторную функцию (вход в моторную кору). В них переключается импульсация от базальных ядер, зубчатого ядра мозжечка, красного ядра среднего мозга. После этого она проецируется в моторную и премоторную кору. Через эти ядра происходит передача в кору сложных двигательных программ, образованных в мозжечке и базальных ядрах.
Ассоциативные ядра таламуса принимают импульсацию не от проводниковых путей сенсорных систем, а от других ядер таламуса. Эфферентные импульсы от этих ядер направляются в ассоциативные поля коры. Кора мозга посылает волокна к ассоциативным ядрам, регулируя их функции. Главной из них является интегративная функция, которая выражается в объединении деятельности как таламических ядер, так и различных зон ассоциативной коры полушарий мозга.
Подушка получает главные входы от коленчатых тел и неспецифических ядер таламуса. Эфферентные импульсы идут от нее в височно – теменно – затылочные зоны коры, участвующих в гностических, речевых и зрительных функциях, а также в восприятии схемы тела.
Медиодорсальное ядро получает входы от гипоталамуса, миндалины, гиппокампа, таламических ядер, центрального серого вещества ствола. Проекция этого ядра распространяется на ассоциативную, лимбическую и лобную кору. Оно участвует в формировании эмоциональной и поведенческой двигательной активности, а также в запоминании.
Латеральные ядра получают зрительную и слуховую импульсацию от коленчатых тел и соматосенсорную импульсацию от вентрального ядра. Интегрированная сенсорная инфа от этих источников далее проецируется в ассоциативную теменную кору и используется в функциях гнозиса, праксиса, формировании схемы тела.
Неспецифические ядра таламуса – более древчяя область, включающая интраламинарную ядерную группу и ретикулярное ядро.
Неспецифические ядра имеют многочисленные входы от других ядер таламуса и внеталамические – эти пути проводят преимущественно болевую и температурную чувствительность. В неспецифические ядра поступает непосредственно или через РФ также часть импульсации по коллатералям от всех специфических сенсорных систем. В неспецифические ядра поступает импульсация из моторных центров ствола (красное ядро, черное вещество), ядер мозжечка, от базальных ядер и гиппокампа, а также от коры большого мозга, особенно лобных долей. Неспецифичные ядра имеют эфферентные выходы на другие таламические ядра, кору больших полушарий, а также нисходящие пути к другим структурам ствола мозга.
Неспецифические ядра вызывают локальную активацию коры больших полушарий в соответствии с корковой проекцией воспринимаемых сенсорных сигналов.
Неспецифические ядра таламуса выступают в роли интегрирующего посредника между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ядрами с другой, объединяя их в единый функциональный комплекс. На кору большого мозга неспецифический таламус оказывает моделирующее влияние.
Гипоталамус включает преоптическую область и область перекреста зрительных нервов, серый бугор и воронку, сосцевидные (мамиллярные тела). В гипоталамусе выделяют 15 – 48 парных ядер, которые разделяют на 3 – 5 групп. Обычно выделяют три основные группы: 1. Передняя группа содержит медиальное преоптическое, супрахиазматическое, перавентрикулярное и переднее гипоталамические ядра; 2) средняя группа включает дорсомедиальное, вентромедиальное, аркуатное (дугообразное) и латеральное гипоталамические ядра; 3. Задняя группа включает супрамамиллярное, премамиллярное, мамиллярные ядра, заднее гипоталамическое и перифорниатное ядра, субталамическое ядро Луиса.
Главные афферентные пути в гипоталамус идут от лимбической степени, коры больших полушарий, базальных ядер и РФ ствола. Основные эфферентные пути гипоталамуса идут в ствол мозга – его РФ, моторные и вегетативные центры, в вегетативные центры спинного мозга, от мамиллярных тел к передним ядрам таламуса и далее в лимбическую систему, от супраоптическрого и паравентрикулярного ядер к нейогипофизу, от вентромедиального и аркуатного ядер к аденогипофизу, а также эфферентные выходы к лобной коре и полосатому телу.
Гипоталамус – важнейший центр интеграции вегетативных, регуляции эндокринной системы, терморегуляции, цикла бодрствование – сон и других суточных биоритмов. Велика его роль в организации поведения, направленного на реализацию биологических потребностей.
Пирамидная система, пирамидный путь, система нервных структур, участвующих в сложной и тонкой координации двигательных актов. У низших позвоночных П. с. нет, она появляется только у млекопитающих, образуя эфферентную часть двигательного анализатора и достигает наибольшего развития у человека. Пирамидный путь начинается преимущественно от пирамидных нейронов сенсомоторной области коры головного мозга. Их длинные отростки (волокна) образуют прямые нисходящие пути к рефлекторным двигательным центрам спинного мозга, по которым осуществляется передача корковой информации. Волокна П. с. спускаются в спинной мозг, не прерываясь; по пути дают ответвления (коллатерали) к ядрам черепных нервов и перекрещиваются (большая часть в продолговатом мозге, меньшая в спинном), переходя на противоположную сторону; далее они проходят в составе передних и боковых столбов спинного мозга, образуя синаптические окончания в каждом его сегменте, передавая импульсы от коры головного мозга его двигательным нейронам непосредственно либо через вставочные нейроны. В составе П. с. человека около 1 млн. нервных волокон. Они делятся в основном на толстые, быстропроводящие (диаметром около 16 мкм, скорость проведения до 80 м/сек) — обеспечивают быстрые фазные движения, и тонкие, медленнопроводящие (диаметром около 4 мкм, скорость проведения от 25 до 7 м/сек), ответственные за тоническое состояние мышц. Повреждения П. с. проявляются параличами, парезами, патологическими рефлексами.
Экстрапирамидная система — совокупность структур головного мозга, участвующих в управлении движениями, поддержании мышечного тонуса и позы, минуя пирамидную систему. Структура расположена в больших полушариях и стволе головного мозга.
Экстрапирамидные проводящие пути образованы нисходящими проекционными нервными волокнами, по происхождению не относящимися к гигантским пирамидным клеткам (клеткам Беца) коры больших полушарий мозга. Эти нервные волокна обеспечивают связи мотонейронов подкорковых структур (мозжечок, базальные ядра, ствол мозга) головного мозга со всеми отделами нервной системы, расположенными дистальнее.
Экстрапирамидная система состоит из следующих структур головного мозга:
базальные ганглии
красное ядро
чёрная субстанция
РФ моста и продолговатого мозга
ядра вестибулярного комплекса
мозжечок
премоторная область коры
полосатое тело
Экстрапирамидная система осуществляет непроизвольную регуляции и координацию движений, регуляцию мышечного тонуса, поддержание позы, организацию двигательных проявлений эмоций (смех, плач). Обеспечивает плавность движений, устанавливает исходную позу для их выполнения.
При поражении экстрапирамидной системы нарушаются двигательные функции (например, могут возникнуть гиперкинезы, паркинсонизм), снижается мышечный тонус.
Экстрапирамидная система объединяет двигательные центры коры головного мозга, его ядра и проводящие пути, которые не проходят через пирамиды продолговатого мозга; осуществляет регуляцию непроизвольных компонентов моторики (мышечного тонуса, координации движений, позы).
Следующее звени экстрапирамидных путей составляют ретикулярно-спинномозговой, красноядерно-спинномозговой, преддверно-спинномозговой и оливоспинномозговой пути, оканчивающиеся в передних столбах и промежуточном сером веществе спинного мозга. Мозжечок включается в экстрапирамидную систему посредством путей, соединяющих его с таламусом, красным ядром и оливными ядрами.
Функционально экстрапирамидная система неотделима от пирамидной системы. Она обеспечивает упорядоченный ход произвольных движений, регулируемых пирамидной системой; регулирует врожденные и приобретенные автоматические двигательные акты, обеспечивает установку мышечного тонуса и поддержание равновесия тела; регулирует сопутствующие движения (например движения рук при ходьбе) и выразительные движения (мимика).
В толще белого вещества полушарий мозга располагаются скопления серого вещества, называемые подкорковыми ядрами (базальные ядра). К ним относятся хвостатое ядро, чечевицеобразное ядро, ограда и миндалевидное тело (рис. 6). Чечевицеобразное ядро, находящееся снаружи от хвостатого ядра, делится на три части. В нем различают скорлупу и два бледных шара.
В функциональном отношении хвостатое ядро и скорлупа объединяются в полосатое тело (стриатум), а бледные шары вместе с черной субстанцией и красными ядрами, расположенными в ножках мозга, – в бледное тело (паллидум). Вместе они представляют очень важное в функциональном отношении образование – стриоппаллидарную систему.
Стриопаллидарная система является важной составной частью двигательной системы. Она входит в состав так называемой пирамидной системы. В двигательной зоне коры головного мозга начинается двигательный – пирамидный – путь, по которому следует приказ выполнить то или иное движение. Экстрапирамидная система, важной составной частью которой является стриопаллидум, включаясь в двигательную пирамидную систему, принимает подсобное участие в обеспечении произвольных движений.
Главным двигательным центром стала кора головного мозга. Стриопаллидарная система стала обеспечивать фон, готовность к совершению движения; на этом фоне осуществляются контролируемые корой головного мозга быстрые, точные, строго дифференцированные движения.
Для совершения движения необходимо, чтобы одни мышцы сократились, а другие расслабились, иначе говоря, нужно точное и согласованное перераспределение мышечного тонуса. Такое перераспределение тонуса мышц как раз и осуществляется стриопаллидарной системой. Эта система обеспечивает наиболее экономное потребление мышечной энергии в процессе выполнения движения. Совершенствование движения в процессе обучения их выполнению (например, отработка до предела отточенного бега пальцев музыканта, взмаха руки косаря, точных движений водителя автомобиля) приводит к постепенной экономизации и автоматизации. Такая возможность обеспечивается стриопаллидарной системой.
Лимбическая система: структуры и их связи, ее роль в регуляции вегетативных функций, в формировании мотиваций и осуществлении эмоциональных и поведенческих приспособительных реакций, в процессах памяти.
Лимбическая система – совокупность структур промежуточного, конечного и среднего мозга, обеспечивающая эмоционально – мотивационные компоненты поведения и интеграцию висцеральных функций организма.
В лимбическую систему включают образования обонятельного мозга – обонятельная луковица и бугорок, гиппокамп, парагиппокампальную, зубчатую и поясную извилины; подкорковые ядра – амигдалу, ядра перегородки; мамиллярные тела гипоталамуса, передние ядра таламуса.
Афферентные входы в лимб систему осуществляются от различных областей головного мозга, а также через гипоталамус от РФ ствола, которая считается главным источником ее возбуждения. Важным стимулирующим афферентным входом являются импульсы от обонятельных рецепторов по волокнам обонятельного нерва, поступающие в различные структуры лимбической системы.
Эфферентные выходы осуществляются через гипоталамус на нижележащие вегетативные и соматические центры ствола и спинного мозга. Другой эфферентный выход проводит возбуждение из лимбической системы в новую кору. Через нее лимбическая система включается в регуляцию высших психических функций.
Кольцевые нейронные связи объединяют различные структуры лимбической системы и дают возможность длительной циркуляции возбуждения, которая является механизмом его пролонгирования, повышения проводимости синапсов и запоминания инфы. Реверберация создает условия для сохранения единого функционального состояния структур замкнутого круга и навязывания этого состояния другим структурам мозга.
Важнейшим циклическим образованием лимбической системы является круг Пейпеца, идущий от гиппокампа через свод к мамилярным телам, от них к передним ядрам таламуса, от него в поясную извилину и от нее через парагиппокампову извилину обратно к гиппокампу. Этот круг имеет большое значение в формировании эмоций, обучении и памяти.
Другой круг (от миндалины к мамилярным телам, от них к лимбической области среднего мозга и обратно к амигдале) имеет огромное значение в формировании агрессивно – оборонительных, пищевых и сексуальных реакций.
Функции лимбической системы. Получая инфу о внешней среде и внутренней среде организма, лимбическая система после сравнения и обработки этой инфы запускает вегетативные реакции и вместе с ассоциативной корой инициирует поведенческие реакции, обеспечивающие адекватное приспособление организма к окружающей среде и сохранению гомеостазиса.
Регуляция висцеральных функций осуществляется преимущественно через гипоталамус. При раздражении лимбической системы повышаются или снижаются секреция различных гормонов аденогипофизом (АКТГ и гонадотропинов), ЧСС, моторика и секреция желудка и кишечника.
В лимбической системе происходит формирование эмоций. Важную роль в этом процессе имеют амигдала и поясная извилина. Электрическая стимуляция амигдалы вызывает отрицательные эмоции - страх, гнев, ярость. Удаление амигдалы приводит к уменьшению агрессивности, повышает тревожность. Нарушается способность оценивать инфу, поступающую из окружающей среды и связывать ее со свои эмоциональным состоянием. Амигдала участвует в процессе сравнивания конкурирующих эмоций, выделения доминирующих эмоций, влияет на выбор поведения.
Амигдала участвует в однократном обучении благодаря свойству индуцировать сильные отрицательные эмоции, что способствует быстрому и прочному формированию временной связи.
Поясная извилина, имеющая многочисленные связи с новой корой и стволовыми центрами, выполняет роль интегратора различных структур мозга, участвующих в формировании эмоций.
Роль лимбической системы в системе обучения связана с лимбическим кругом Пейпеца. Главную роль играют гиппокамп и связанные с ним задние зоны лобной коры. Их деятельность необходима для консолидации памяти – перехода кратковременной памяти в долговременную. Повреждение гиппокампа ведет к резкому нарушению усвоения новой инфы, образования промежуточной и долговременно памяти.
Электрофизиологической особенность. Гиппокампа является то, что в ответ на раздражение, стимуляцию РФ и заднего гипоталамуса в нем развивается синхронизация электрической активности в виде тета – ритма. При этом в новой коре возникает десинхронизация виде бета – ритма. Другая особенность - способность на стимуляцию отвечать длительным увеличением амплитуды возбуждающих постсинаптических потенциалов, что приводит к синаптической потенциации и запоминанию инфы. При этом на дендритах пирамидных нейронов гиппокампа увеличивается число шипиков, что свидетельствует об усилении синаптической передачи возбуждающих и тормозных влияний через аксоношипиковые синапсы.
Сенсорная функция. В лимбической системе находится корковый отдел системы обоняния (парагиппокампальная извилина, крючок, гиппокамп). Ее главный афферентный выход через свод, мамиллярные тела, передние ядра гипоталамуса на другие структуры лимбической системы и далее на висцеральные органы объясняет наличие резко выраженного эмоционального компонента в обонятельном восприятии и возможность лечебного воздействия пахучих веществ.
Функции новой коры, функциональное значение первой и второй соматосенсорных зон, моторные зоны коры больших полушарий (их локализация и функциональное значение). Полифункциональность корковых областей, функциональная пластичность коры.
Новая кора – слой серого вещества общей площадью 1100 – 1900 см2, покрывающий около 71 % площади больших полушарий. Она осуществляет высшую регуляцию функций организма, обеспечивает психическую деятельность и поведение.
Свойство многофункциональности позволяет корковой структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную функцию. В основе мультифункциональности лежат: многоканальность поступления в кору мозга афферентного возбуждения, перекрытия афферентных возбуждений особенно на таламическом и корковом уровнях, модулирующее влияние различных структур на корковые функции, взаимодействие корково – подкорковых и межкорковых путей проведения возбуждения.
Функционально кору разделяют на сенсорную, ассоциативную, двигательную и лимбическую области.
Сенсорные области – зоны, в которые проецируется информация от сенсорных раздражителей. Они расположены в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от специфических сенсорных ядер таламуса.
Зоны коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма, называют первичными сенсорными зонами. Они состоят преимущественно из моносенсорных нейронов и формируют ощущения одной модальности. В первичных сенсорных зонах имеется четкое пространственное представительство частей тела, их рецептивных полей. Нейроны первичных зон имеют наименьший порог возбуждения. Вокруг первичных сенсорных зон находятся вторичные сенсорные поля. Нейроны этих полей являются преимущественно бисенсорными.
Кору постцентральной извилины темненной доли и соответствующую ей часть парацентральной дольки на медиальной поверхности полушарий обозначают как первичную соматосенсорную область. В ней представлена древняя топографическая карта распределения кожных рецепторов. В этой области имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности от мышечных, суставных, сухожильных рецепторов.
Вторичная соматосенсорная зона расположена в стенке боковой борозды, на границе ее пересечения с центральной бороздой.
Ассоциативные области коры включают участки новой коры большого мозга, которые выполняют интегрирующую роль между сенсорными и двигательными участками.
Полисенсорность нейронов ассоциативной коры создается ее кортико – кортикальными связями с сенсорными зонами, так и главным ее афферентным входом от ассоциативных ядер таламуса, в которых уже произошла обработка инфы от разных чувствительных путей.
Формирование программ целенаправленного поведения происходит в лобной ассоциативной коре. Реализация этой функции основывается на других функциях этой системы: 1. Формирование доминирующей мотивации, определяющей направление поведения человека; 2. Обеспечении вероятностного прогнозирования, что выражается изменением поведения в ответ на изменение обстановки окружающей среды и доминирующей мотивации; 3. Самоконтроле действий путем постоянного сравнения результата действия с исходными намерениями, что связано с созданием аппарата поведения (акцептор результата действия).
Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, моторную кору и ядра экстрапирамидной системы. Основными функциями является гнозис, практис и формирование схемы тела.
В двигательной коре выделяют первичную и вторичную моторные зоны.
В первичной двигательной коре (прецентральная извилина, поле 4) расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела. Двигательные реакции на раздражение первичной коры осуществляются с минимальным порогом, они представлены элементарными сокращениями мышц противоположной стороны тела.
Вторичная моторная кора расположена в лобных долях на латеральной стороне полушарий, впереди прецентральной извилины (премоторная кора) и на медиальной стороне полушария (дополнительная моторная кора). Она осуществляет высшие двигательные функции, связанны с программированием и координацией произвольных движений. Кора поля 6 получает основную часть импульсации от базальных ядер и мозжечка и участвует в перекодировании инфы о программе сложных движений. При раздражении коры поля 6 возникают сложные координированные движения. В премоторной коре расположен центр моторной речи Брока. Там же расположен музыкальный моторный центр, определяющий тональность речи, способность петь.
Вегетативная нервная система: функциональные отличия от соматической нервной системы, особенности симпатического и парасимпатического отделов, синергизм и относительный антагонизм - их влияния, механизм передачи возбуждений в вегетативных ганглиях и на рабочий орган (виды рецепторов, медиаторы).
ВНС – часть НС, регулирующая работу внутренних органов, просвет сосудов, обмен веществ и энергии, обеспечивая гомеостазис организма.
Основные отличия ВНС от соматической нервной системы.
Функцией ВНС является поддержание постоянства внутренней среды, приспособление ее к изменяющимся условиям окр среды и деятельности орг – ма.
Влияние ВНС на организм не находятся под непосредственным контролем сознания.
Регуляция функций внутренних органов может осуществляться, хотя и менее совершенно, при полном нарушении связи с ЦНС. Объясняется это тем, что эффекторный нейрон ВНС, находится за пределами ЦНС: либо в экстра либо в интраорганных вегетативных ганглиях, что обеспечивает формирование периферических рефлекторных дуг.
Генерализованный (диффузный) характер распространения возбуждения в периферическом отделе ВНС. Объясняется это, во – первых, феноменом мультипликации вегетативных ганглия, во – вторых, тем, что большинство адренергических нейронов обладает длинными постганглионарными тонкими аксонами, многократно ветвящимися в органах образующими адренергические сплетения.
Низкая скорость проведения возбуждения в вегетативных нервах.
Низкая лабильность нейронов вегетативных ганглиев. Она объясняется длительной синаптической задержкой и продолжительной следовой гиперполяризацией нейрона.
Выделяют два отдела ВНС: симпатический и парасимпатический. Различие этих отделов заключается в том, что у эффекторных нейронов, а соответственно постганглионарных волокон различные медиаторы: у парасимпатической – АХ, у симпатической – норадреналин.
Преганглионарные волокна и симпатической и парасимпатической НС являются холинергическими.
Симпатическая НС.
Преганглионарные нейроны СНС расположены в боковых рогах, сегменты С8 – L2 – центр Якобсона. Аксоны преганглионарных нейронов синаптически связаны с нейронами симпатических ганглиев, расположенных экстраорганно. От нейронов симпатических ганглиев идут постганглионарные волокна, иннервирующие все органы, сосуды и ткани организма. В последнем шейном и двух первых грудных сегментах спинного мозга расположен спиноцилиарный центр, иннервирующий гладкие мышцы глаза, в том числе мышцу, расширяющую зрачок. Эффекторные нейроны этого пути лежат в краниальном шейном симпатическом ганглии. От 2 – 4 грудного сегмента спинного мозга отходят симпатические волокна, иннервирующие слюнные железы. В Th1 – Th5 расположены преганглионарные симпатические волокна, иннервирующие сердце.
Медиатором преганглионарных симпатических волокон является АХ, он активирует Н – холинорецепторы. В симпатических ганглиях высвобождаются нейропептиды, выполняющие модулирующую функцию. Медиатором постганглионарных симпатических волокон являются катехоламины – норадреналин, адреналин, дофамин. Синтезируется норадреналин из аминокислоты тирозина, частично в терминальных симпатических окончания, частично в теле нейрона. Катехоламины высвобождаются при возбуждении нервного окончания путем везикулярного экзоцитоза, реализуемого с помощью кальция.
Инактивирование медиатора, выделившегося в синаптическую щель, осуществляется за счет обратного его захвата пресинаптической терминалью, часть разрушается катехол – о –метилтрансферазой. Часть норадреналина диффундирует в интерстиций и кровь.
Характеристика циторецепторов.
Большинство циторецепторов связано с G – белками, представляющими собой ГТФ – связывающие белки. Все циторецепторы делятся на два класса: ионотропные и метаботропные. Действие медиатора на ионотропные рецепторы активирует их ионные каналы с помощью G – белка без вторых посредников. Активация ионных каналов нейромедиаторами посредством метаботропных рецепторов реализуется с помощью вторых посредников.
При передаче сигнала от нервных волокон посредством и ионотропных и метаботропных рецепторов к другим клеткам в последних вслед за электрофизиологическими активируются биохимические процессы. Это реосинтез АТФ. В нервных клетках запускаются биохимические процессы, обеспечивающие хранение инфы.
Вторыми посредниками альфа 1 адренорецепторов является ИФ3 (инизитол 3 фосфат); альфа два и бета адренорецепторов – система адениатциклаза – цАМФ.
При избытке медиатора развивается снижение чувствительности клетки к медиатору (десенситизация), а при недостатке медиатора повышение их чувствительности (сенситизация).
Преганглиоанарые симпатические волокна образуют синапсы на клетках мозгового слоя надпочечников. При возбуждении этих волокон из надпочечников выбрасывается смесь из адреналина и норадерналина.
Парасимпатическая НС.
Парасимпатическую иннервацию не получают: кровеносные сосуды, органы чувств и мозговое вещество надпочечников, скелетные мышцы. Парасимпатическая система способствует поддержанию гомеостаза.
Парсимпатические нервные волокна проходят в составе черепных нервов (3 пера – мезенцефальный отдел; 7, 9 и 10 пары – бульбарный отдел) и тазовых нервов – сакральный отдел спинного мозга (S2 – S4). Последние иннервируют нисходящую часть ободочной кишки и тазовые органы. Парасимпатические Волокна третьей пары иннервирую глазные мышцы, регулируя диаметр зрачка и степень аккомодации. Парасимпатические веточки 7 пары иннервируют слизистую оболочку носа, неба, слезную железу. Парасимпатические секреторные волокна 9 пары подходят к околоушной железе. Парасимпатические ганглии расположены внутри органов, а в области головы и таза вблизи органов. От нервных клеток парасимпатических ганглиев идут короткие постганглионарные парасимпатические волокна, иннервирующие эффекторные клетки органов.
Медиатором преганглионарных волокон является АХ. Он действует на ионотропные N – холинорецепторы постсинаптической мембраны ганглионарного нейрона. Медиатором постганглионарных парасимпатических волокон также является АХ. Часть АХ, высвобождаемого терминалями разрушается ацетилхолинэстеразой, с образованием холина и уксусной кислоты, которые захватываются пресинаптической мембраной и вновь используются для синтеза АХ, часть диффундирует в интерстиций.
Постганглионарные парасимпатические терминали образуют синапсы с эффекторными рецепторами, которыми являются М – холинорецепторы.
Влияние парасимпатической и симпатической систем обычно разнонаправлены – это основа взаимодействия. Оба отдела ВНС действуют синергично, обеспечивая получение для организма приспособительного полезного результата.
Роль различных отделов ЦНС в регуляции вегетативных функций: спинной, продолговатый и промежуточный мозг, лимбическая система, мозжечок, кора больших полушарий. Афферентное звено вегетативной нервной системы и его значение в регуляции функций внутренних органов.
Гипоталамические структуры предопределяют качество вегетативного обеспечения той или иной конкретной соматической деятельности организма, приводя режимы работы вегетативных эффекторов и параметры обмена веществ в соответствие с текущими потребностями организма.
Рецепторные нейроны гипоталамуса практически не адаптируются: они генерируют импульсы до тех пор, пока показатель не нормализуется в результате адаптивного изменения работы вегетативных эффекторов за счет эфферентных влияний гипоталамуса. Воздействие гипоталамуса на вегетативные эффекторы осуществляется посредством симпатических, парасимпатических и серотонинергических влияний, а также через эндокринные железы.
Роль различных групп ядер гипоталамуса определяется их связью с симпатическими или парасимпатическими отделами ВНС. Раздражение передних ядер гипоталамуса вызывает в организме изменения, подобные тем, что возникают при активации парасимпатической НС. Раздражение задних ядер сопровождается эффектами, наблюдаемыми при стимуляции симпатической НС. В гипоталамусе локализуются центры регуляции цикла сон – бодрствование.
Лимбическая система (в состав входят: гипоталамус, гиппокамп, грушевидная извилина, периамигдалярная кора, поясная извилина, область перегородки, миндалевидный комплекс, таламус, лимбическая зона среднего мозга). Раздражение отдельных ее структур может вызвать изменения деятельности ССС, пищеварительной и других систем. Это влияние осуществляется при помощи ВНС и опосредованно через эндокринные железы.
РФ управляет вегетативными функциями посредством активации симпатической НС, ее нейроны формируют жизненно важные центры продолговатого мозга – дыхательный и сердечно – сосудистый. Таламус ограничивает поступление афферентной импульсации в кору большого мозга от внутренних органов, обеспечивая ее большую активность в регуляции соматических функций. Мозжечок с помощью симпатической Нс принимает участие в регуляции функций внутренних органов. Мозжечок стабилизует гомеостазис.
Кора большого мозга – высший интегративный центр регуляции всех функций, в том числе вегетативных. Раздражение различных зон коры вызывает изменение деятельности всех органов, имеющих вегетативную иннервацию.
Афферентное звено ВНС.
Периферические афферентные пути: 1. В составе блуждающего нерва содержится до 90 % афферентных волокон, тела нейронов этих волокон находятся в узловатом ганглии. 2. В тазовом нерве афферентные волокна составляют 50 %. 3. В составе симпатических стволов также имеются афферентные волокна, в совокупности они составляют 50 % от всего количества волокон. Тела нейронов, от которых такие волокна, локализуются в интра и экстраорганных превертебральных ганглиях. Часть этих афферентных волокон образуют синапсы на вставочных клетках экстарорганных ганглиев, формируя афферентное звено периферической экстраорганной вегетативной рефлекторной дуги. Часть афферентных волокон симпатических нервов идет в спинной мозг в составе задних корешков, вместе с чувствительными соматическими волокнами. 4. Рецептивные поля внутренних органов снабжены чувствительными волокнами, являющимися отростками афферентных нейронов межпозвоночных спинальных ганглиев и гомологичных им черепных нервов.
Сенсорные рецепторы афферентов ВНС воспринимают различные изменения внутренней среды, все они являются первичными рецепторами: механо, хемо, термо, осмо.
Афферентные пути в ЦНС. Афферентные волокна ВНС, вступив в спинной мозг, образуют синапсы с промежуточными нейронами данного сегмента, соседних или вышерасположенных сегментов, вступают в продолговатый мозг, где находится второй афферентный нейрон, посылающий свой аксон в зрительный бугор к третьему афферентному нейрону. Аксон которого направляется в соматосенсорную зону коры большого мозга.
ИНТЕГРАТИВНАЯ ДЕЯТЕЛЬНОСТЬ ОРГАНИЗМА
АНАЛИЗАТОРЫ (СЕНСОРНЫЕ СИСТЕМЫ)
Анализаторы (И.П. Павлов): понятие, классификация анализаторов, три отдела анализаторов и их значение, понятие об органах чувств, значение внешних анализаторов.
Анализаторы = сенсорные системы – совокупность центральных и периферических образований НС, обеспечивающих восприятие и анализ раздражителей и участвующих в формировании ответной реакции.
Классификация анализаторов.
Сенсорные системы внешней среды. Это системы зрения, слуха, тактильная, температурная, обоняния, вкуса, возбуждение которых воспринимается субъективно в виде ощущения. Роль этих систем – познание внешнего мира, приспособление к окружающей среде, поддержание тонуса ЦНС.
Сенсорные системы внутренней среды воспринимают и анализируют изменение различных показателей деятельности внутренних органов. Возбуждение этих систем обычно не ощущается.
Значение – участие в регуляции функций внутренних органов, поддержании гомеостазиса, обеспечение приспособления организма к различным условиям его жизнедеятельности.
Имеется несколько внутренних сенсорных систем 6 химизма внутренней среды, осмотического давления, количества жидкости, наполнения внутренних органов.
Сенсорные системы положения тела и его частей. Это вестибулярная и проприоцептивная системы. Роль заключается в регуляции мышечного тонуса и поддержании с помощью этого естественной позы, в восстановлении нарушенной позы, в координации движений. Активация этих систем ощущается при участии сознания.
Система боли информирует о повреждающих действиях на организм и патологических процессах.
Структурно – функциональная организация сенсорных систем.
Анализаторы имеют три отдела: периферический, проводниковый и центральный.
Периферический отдел – рецептор. Назначение рецептора – восприятие раздражителей и первичный их анализ. Восприятие раздражителей осуществляется посредством трансформации энергии раздражителя в рецепторный потенциал, инициирующий возникновение в афферентном нервном окончании нервного импульса.
Проводниковый отдел включает совокупность всех нервных элементов, по которым сигнал от рецепторов достигает коры большого мозга: афферентные (периферические) и промежуточные нейроны спинного мозга, стволовых и подкорковых структур ЦНС.
В проводниковом отделе выделяют специфический (проекционный) и неспецифический пути. Специфический путь обеспечивает формирование ощущений определенной модальности. От рецептора импульсы идут по строго обозначенным для данного анализаторв специфическим путям с переключением на различных уровнях ЦНС.
Неспецифический путь формируется ретикулярной формацией. На уровне ствола мозга от специфических путей отходят коллатерали к ретикулярной формации, на которых могут конвергировать афферентные возбуждения различной модальности, способствуя взаимодействию сенсорных систем. Возбуждение по неспецифическим путям проводится медленно, через большое число синапсов.
Центральный отдел сенсорной системы. В корковом отделе выделяют первичную зону, куда поступают импульсы от рецепторов одного вида и вызывают ощущения одной модальности, и вторичную зону, расположенную в близости от первичной. Нейроны вторичной зоны преимущественно бисенсорные, возбуждаются при действии двух раздражителей. Сенсорные зоны коры перекрывают друг друга, что обеспечивает взаимодействие различных сенсорных систем и процесс компенсации нарушения функций.
Взаимодействие различных зон коры обеспечивает формирование соответствующих ощущений и конечный анализ и синтез поступающей в ЦНС информации.
Органы чувств – периферические образования, воспринимающие и частично анализирующие раздражители окружающей среды.
Свойства анализаторов и приспособление организма в окружающей среде, критерии для оценки чувствительности анализаторов к адекватным раздражителям.
Высокая чувствительность сенсорных систем к адекватному раздражителю – обеспечение восприятия очень слабых раздражителей.
Способность к адаптации сенсорных систем к постоянной силе длительно действующего раздражителя. Адаптация в основном заключается в понижении абсолютной и повышении дифференциальной чувствительности.
Сенситизация – повышение чувствительности сенсорных систем может наблюдаться при возбуждении симпатической нервной системы, выбросе катехоламинов надпочечниками, при увеличении тироксина в крови, в условиях действия слабых раздражителей.
Инерционность сенсорных систем – отставание возникновения ощущения от начала действия раздражителя и продолжение ощущений некоторый период после его выключения.
Системный механизм узнавания и восприятия.
Критерии оценки чувствительности сенсорных систем.
Порог ощущения (абсолютный порог) – минимальная сила раздражения, вызывающая такое возбуждение сенсорной системы, которое воспринимается субъективно в виде ощущения.
Порог различения – минимальное изменение силы действующего раздражителя, воспринимаемое субъективно в виде изменения интенсивности ощущения.
Порог пространства – минимальное расстояние между двумя раздражаемыми точками, при котором раздражения воспринимаются как раздельные.
Интенсивность ощущений.
Кодирование информации в анализаторах: понятие о кодировании, коды нервной системы, кодирование в рецепторах (кодируемые характеристики раздражителей, механизм), кодирование в проводниковом и в корковом (центральном) отделах анализаторов. Механизм узнавание предметов и явлений, формирование, образов впервые наблюдаемых предметов.
Кодирование - процесс преобразования информации в условную форму, удобную для передачи по каналу связи.
Универсальным кодом является ПД, который распространяется по нервным волокнам. Передача сигналов от одной клетки к другой во всех отделах сенсорных систем осуществляется при помощи химического кода – медиатора.
В сенсорных системах кодируются качественная характеристика раздражителя (его вид), сила раздражителя, время его действия, место действия раздражителя на организм и локализацию его в окружающей среде.
Кодирование вида раздражителя осуществляется за счет специфичности рецепторов, что и позволяет организму отличать один вид раздражителя от другого.
Сила раздражителя кодируется величиной РП, и, следовательно, изменением частоты импульсов, возникающих в окончании чувствительного нервного волокна при изменении силы действующего раздражителя.
Локализация действия раздражителя кодируется тем, что возбуждаются только те рецепторы, на которые действуют раздражители, при этом рецепторы различных участков тела посылают импульсы в определенные зоны коры. Пространство на теле организма также кодируется величиной площади, на которой возбуждаются рецепторы.
Кодирование времени действия раздражителя на рецепторы состоит в том, что они начинают возбуждаться с началом действия раздражителя на рецепторы и прекращают возбуждаться сразу после выключения раздражителя.
Кодирование в проводниковом отделе СС.
Кодирование в проводниковом отделе анализатора осуществляется на «станциях переключения», то есть при передаче сигнала от одного нейрона к другому, где происходит смена кода. В нервных волокнах информация не кодируется, они исполняют роль проводов, по которым передается информация, закодированная в рецепторах или поступающая от различных нейронов в ЦНС к другим клеткам. Импульсы в отдельном нервном волокне формируются в пакеты – паттерны, – между которыми могут быть различные интервалы, в паттернах – различное число импульсов, между отдельными паттернами – различные интервалы. В нервном стволе, состоящем из множества волокон, может изменяться количество возбужденных волокон – это зависит от количества возбужденных рецепторов или нейронов на предыдущем звене следования сигнала и при переходе его с одного нейрона на другой. На «станциях переключения», например, в таламусе, информация кодируется, во-первых, за счет изменения объема импульсации на входе и выходе, а во-вторых, за счет пространственного кодирования, то есть связи определенных нейронов с определенными рецепторами. По мере поступления импульсов к вышележащим отделам ЦНС уменьшается частота разрядов нейронов и длительная импульсация превращается в короткие пакеты импульсов. Продолжительность разряда большинства нейронов уже не соответствует длительности стимула. В таламусе начинают формироваться ощущения, что является началом субъективного анализа и синтеза действующих на организм раздражителей. В проводниковом отделе анализатора имеются нейроны, возбуждающиеся не только при появлении стимула, но и при его выключении, что связано с активностью рецепторов и результатом взаимодействия самих нейронов, а также нейроны, которые частично отражают свойства стимула (нейроны-детекторы). Количество нейронов-детекторов возрастает на каждом последующем уровне анализатора, поэтому структура ответа в нейронах не соответствует структуре ответа в рецепторе. В то же время на каждом последующем уровне анализатора имеются нейроны, которые дублируют свойства нейронов предыдущего отдела, что создает основу надежности функции анализатора. Наряду с возбуждением в сенсорных ядрах происходит и торможение. Тормозные процессы осуществляют фильтрацию и дифференциацию сенсорной информации. Они обеспечивают также контроль сенсорной информации, тем самым устраняют избыточные несущественные сигналы. Такой механизм реализуется за счет латерального и возвратного торможения в процессе восходящих и нисходящих влияний. Таким образом, вследствие переработки информации на «станциях переключения» проводникового отдела анализатора в корковый отдел поступает сильно измененная в количественном и качественном отношении импульсация. Нейроны проводникового отдела анализатора распознают вид раздражителя и структуру ответа, зависящую от характера раздражителя.
Кодирование в корковом отделе СС.
В основе лежат элекрофизиологические процессы, вызывающие в нейронах нейрохимические реакции, а те в свою очередь обусловливают структурные изменения в нейронах – информация кодируется с помощью синтеза белка.
Кодирование с помощью элекрофизиологических процессов является пространственно – временным: нервные импульсы вначале поступают в первичную сенсорную зону, где локализуются моносенсорные нейроны. Затем импульсация поступает через ассоциативные ядра таламуса во вторичную сенсорную зону, где локализуются преимущественно бисенсорные нейроны. Позже возбуждение поступает в ассоциативную кору, где локализуются полисенсорные нейроны.
Зрительный анализатор: назначение основных структурных элементов глаза: механизм приспособления глаза к ясному видению в условиях изменения расстояния до объекта, при перемещении предмета или появления его в новом участке поля зрения, при фиксации взора на неподвижном предмете.
Орган зрения (глаз) – парное, подвижное сферическое образование, включающее рецепторы, часть проводникового отдела и вспомогательные структуры.
Световоспринимающий аппарат (фоторецепторы глаза) обеспечивает восприятие квантов света – кодирование поступающей информации и передачу ее по афферентным путям (волокнам зрительного нерва) в ЦНС.
Оптическая система (слезная железа, роговица, водянистая влага, хрусталик, стекловидное тело) предназначена для фокусирования световых лучей на сетчатке и формирования в области ее центральной ямки перевернутого, уменьшенного изображения рассматриваемых объектов и явлений.
Глазодвигательный аппарат (наружные мышцы глазного яблока – 4 прямые и 2 косые, подниматель верхнего века и орбитальная мышца, внутренние мышцы глаза) производит необходимую для зрения фиксацию глаз, их поворот, и установку зрительных осей, а также аккомодацию зрения.
Система жизнеобеспечения структур глаза поддерживает кровоснабжение, выработку внутриглазной жидкости, регуляцию гидро и гемодинамики.
Рецепторный отдел системы зрения представлен четырьмя видами фоторецепторов – палочки и три вида колбочек, расположенные в сетчатке. Рецепторный слой прилежит к слою клеток пигментного эпителия. В каждом глазу насчитывается около 110 – 125 млн. палочек и 6 -7 млн. колбочек. Колбочки плотно упакованы в центральной ямке (место наилучшего зрения), что обеспечивает высокую остроту зрения. Палочки распределены на периферии сетчатки, обусловливают высокую световую чувствительность при сумеречном зрении. Плотность распределения палочек уменьшается по мере удаления от центра сетчатки. Клеточные элементы в сетчатке устойчиво фиксированы благодаря наличию между ними опорных глиальных элементов: астроцитов, волокон Мюллера.
Фоторецепторные клетки состоят из двух сегментов: наружного, содержащего светочувствительный пигмент, и внутреннего, включающего митохондрии, рибосомы и аппарат Гольджи. Наружный сегмент, погруженный в слой пигментного эпителия сетчатки, поглощает кванты света и преобразует их энергию в рецепторный потенциал.
Палочки содержат пигмент родопсин, поглощающий электромагнитное изучение. Пороговая чувствительность палочек составляет 2 – 12 квантов света. Палочки обеспечивают периферическое зрение, сумеречное черно – белое зрение, восприятие движений.
Каждая колбочка содержит по одному из трех пигментов – сине – голубой, зеленый, красный. Порог чувствительности разных колбочек 30 – 110 квантов. Колбочки обеспечивают зрение в условиях хорошей освещенности, высокую остроту зрения, цветовосприятие.
Механизм рассмотрения разноудаленных предметов.
Аккомодация заключается в сохранении фокусировки изображения рассматриваемого объекта на сетчатке за счет изменения кривизны хрусталика и его преломляющей силы.
Хрусталик – прозрачное эластичное бессосудистое тело в форме двояковыпуклой линзы.
Хрусталик заключен в капсулу, которая по краям переходит в фиксирующую его циннову связку, соединенную с ресничной мышцей. При сокращении ресничной мышцы натяжение цинновой связки уменьшается и хрусталик становится более выпуклым. Преломляющая сила хрусталика увеличивается, глаз настраивается на рассматривание близких предметов. Когда человек смотрит вдаль, ресничная мышца расслаблена, циннова связка натянута, что приводит к растяжению капсул хрусталика, его уплощению и снижению преломляющей силы.
Цилиарная мышца, от которой зависит кривизна хрусталика, иннервируется волокнами парасимпатической ветви 3 пары черепных нервов. Стимуляция М3 – холинорецепторов цилиарных мышц АХ вызывает их сокращение. Адекватным стимулом для изменения степени аккомодации является нечеткость изображения на сетчатке (ретинальный рефлекс).
Конвергентные и дивергентные движения глазных яблок – соответственно сведение или разведение зрительных осей глаз для обеспечения удерживания объекта в центре желтого пятна сетчатки.
Изменение размеров зрачка. Реакция зрачков происходит синхронно с движением глаз. При рассматривании близко расположенных объектов наряду со сведением (конвергенцией) зрительных осей происходит сужение зрачков – конвергентная реакция. Она запускается рефлекторно от сокращающихся прямых мышц глаза и уменьшает искажения изображения на сетчатке, вызываемого сферической абербацией. Последняя заключается в том, что периферические части хрусталика сильнее преломляют лучи, поэтому изображение на сетчатке при большом диаметре зрачка становится менее четким.
Бинокулярное зрение. Формирование бинокулярного зрения обеспечивается функционированием механизма фиксации взора, синхронизации движений обоих глаз, удерживания изображения в области центральной ямки сетчатки. Относительная диспарантность (расхождение) – небольшое различие в изображении объектов на корреспондирующих точках обоих глаз. Единый объемный бинокулярный образ объекта возникает в мозге человека в результате слияния сетчаточных изображений (фузий) и интегрирования информации, поступающей от корреспондирующих участков сетчатки обоих глаз в корковых отделах зрения.
Приспособления глаза для рассматривания движущихся и неподвижных объектов. Ясному видению движущегося объекта способствуют содружественные движения глаз.
Произвольные движения глаз со скоростью движения объекта влево, вправо, вверх, вниз, осуществляются благодаря содружественной деятельности глазодвигательных мышц.
Плавные, непроизвольные следящие движения глаз обеспечивают совмещение изображения на сетчатке с центральной ямкой. Если изображение смещается на край ямки, срабатывает фиксационный рефлекс – непроизвольные мелкие скачкообразные быстрые движения (саккады), обеспечивающие коррекцию совмещения изображения на сетчатке с центр ямкой.
При оценке неподвижного объекта и фиксации взора предотвращается адаптация фоторецепторов с помощью быстрых непроизвольных постоянных содружественных движений глаз (тремор), совершаемых с большой с скоростью и превращающих постоянный раздражитель в прерывистый, т.к. световые лучи быстро перемещаются от одних рецепторов к др, поэтому не успевает произойти адаптация. Глаза совершают также постоянные медленные движения (медленный дрейф). При этом в случае фиксации взора и попадании изображения вследствие дрейфа на край центральной ямки срабатывает фиксациооный рефлекс (саккады).
При смещении взора с одной детали неподвижного объекта на др используются плавные произвольные движения глаз.
Управление наружными мышцами глаз осуществляется альфа мотонейронами ядер 3, 4, 6 пары черепных нервов, являющихся общим конечным путем для всех надъядерных центров головного мозга, принимающих активное участие в регуляции движений глаз. Активность мотонейронов глазодвигательных ядер контролируется нейронами центров горизонтального (варолиев мост) и вертикального (средний мозг) взоров. Рефлекторные и произвольные движения глаз находятся под контролем центров, расположенных в различных областях мозга – зрительной, теменной, лобной областях коры, в претектальной зоне, верхних бугорках четверохолмия, вестибулярных ядрах, мозжечке.
Механизм приспособления глаза к ясному видению в условиях различной освещенности, при рассматривании крупных объектов и их деталей, при изменении длины световой волны (цветовое зрение). Аномалии рефракции глаза. Проводниковый и корковый отделы зрительного анализатора.
Механизм обеспечения ясного зрения при изменениях освещенности. Главные из них темновая и световая адаптации. Они осуществляются с помощью фотохимических процессов – расщепление зрительных пигментов в колбочках и палочках на свету и ресинтеза их в темноте. Во время темновой адаптации увеличиваются функциональные размеры рецептивных полей биполярных и ганглиозных клеток. ЦНС контролирует эти процессы через регуляцию активности нейронов сетчатки.
Защитный рефлекс смыкания век осуществляется при воздействии сверхяркиих стимулов и внезапной сильной засветке сетчатки. Структурами рефлекса являются связи сетчатки с подкорковыми зрительными центрами и мотонейронами лицевого нерва, иннервирующими круговую мышцу глаза.
Зрачковый рефлекс позволяет изменять в 30 раз интенсивность светового потока, попадающего на сетчатку, путем изменения диаметра зрачка. При увеличении освещенности и рассматривании близко расположенных предметов происходит сужение зрачка (миоз) за счет сокращения мышечного сфинктера по механизму для цилиарной мышцы. При уменьшении освещенности, психическом возбуждении и эмоциональных реакциях, связанных с повышением тонуса симпатического отдела нервной системы, зрачок расширяется (мидриаз) за счет сокращения мышцы – дилататора, вызываемого норадреналином через альфа 1 адренорецепторы миоцитов. Дилататор получает симпатическую иннервацию о нейронов спинного мозга через верхний шейный симпатический узел. Регуляция функции дилататора происходит с помощью супрануклеарного гипоталамического центра, который посредством РФ с симпатическим цилиоспинальным центром, расположенным в сегментах C8 – Th2.
Восприятие крупных отделов и их деталей. Зрительное восприятие таких объектов обеспечивается за счет центрального и периферического зрения, а также произвольного движения глаз при переводе взгляда с одной точки объекта на др.
Наиболее тонкая оценка деталей предмета возможна в том случае если его изображение попадает в центральную ямку, обеспечивающую наибольшую остроту зрения.
Периферическое зрение характеризуется величиной поле зрения. Это совокупность точек углового пространства, одновременно воспринимаемых при фиксируемом взоре.
Цветовое (хроматическое зрение) – способность зрительного аппарата дифференцировать электромагнитные световые волны по их длине с формированием ощущения света.
Ощущение света связано с освещенностью, по мере ее уменьшения престают различаться красные цвета, позднее синие и фиолетовые.
В сетчатке имеется три вида колбочек, комбинация возбуждения разных колбочек вызывает ощущения цвета. Трихромазия – наличие трех видов колбочек для нормального цветовосприятия. Отсутствие колбочек сопровождается моно или ахромазией (полная цветовая слепота), при недостатке одного вида возникает частичная цветовая слепота (цветоаномалия). Протанопия – слепота на красный цвет, дейтеранопия – зеленого (самая частая), тританопия – слепота на фиолетовый и дефектное зрение на синий и желтый.
В соответствии с цветооппонентной теорией Геринга существуют специфические анатомические нейронные механизмы, связывающие попарно красный и зеленый, желтый и синий, черный и белый. Цветовые ощущение связаны не только со спектральным составом излучений, но и с особенностями организации рецептивных полей биполяров и ганглиозных клеток, их включением/выключением при попадании световых волн в центр или на периферию рецептивного поля и в конечном итоге субъективным формированием оттенков цвета в зрительной коре.
Рефракция – суммарная преломляющая сила оптического аппарата глаза. При нормальной рефракции (эмметропия) лучи от далеко расположенных предметов после прохождения через оптический аппарат фокусируются в центральной ямке сетчатки.
Миопия (близорукость) – лучи фокусируются впереди сетчатки из – за повышенной суммарной преломляющей силы глаза или большой длины глазного яблока. Очки с двояковогнутыми линзами.
Гиперметропия (дальнозоркость) – лучи в силу слабой преломляющей способностью глаза или при малой длине глазного яблока фокусируются за сетчаткой. Двояковыпуклые линзы.
Астигматизм – нарушение рефракции, обусловленное различной роговицы или хрусталика в разных меридианах. Световой пучок невозможно собрать. Цилиндрические линзы.
Проводниковый отдел системы зрения. На уровне нейронов сетчатки обеспечиваются передача и обработка зрительных сигналов от фоторецепторов. Эти процессы начинаются на уровне синапсов фоторецепторов с биполярными и горизонтальными клетками и идут к сетчатке в двух взаимосвязанных направлениях – горизонтальном и вертикальном. Горизонтальное направление (фоторецепторы – горизонтальные клетки) обработки сигналов представлено в основном на периферии сетчатки и обеспечивают суммирование сигналов, восприятие слабых сигналов и движений. Вертикальное направление (фоторецептор – биполярная клетка – ганглиозная клетка) представлено в центральной ямке; обеспечивает высокую разрешающую способность зрения за счет передачи сигналов с одного рецептора на одну ганглиозную клетку. На уровне сетчатки осуществляются обработка и передача зрительных сигналов, в которых участвуют многие функциональные типы нейронов, используются различные нейромедиаторы, обеспечивающие взаимные возбуждающие и тормозные влияния.
On – биполярные клетки активируются на свету, off – клетки в темноте, что обеспечивает передачу инфы в ЦНС по двум независимым нейронным каналам.
Биполярные клетки передают сигналы на ганглиозные клетки, где впервые возникают ПД (2 – й вертикальный нейрон), а также на амакриновые клетки.
ПД ганглиозных клеток интегрируются и передаются в ЦНС в виде нервных импульсов по аксонам ганглиозных клеток , формирующим зрительный нерв.
Ганглиозные клетки – многоотростчатые нейроны, образующие своими аксонами зрительный нерв, реагируют на множество свойств зрительного объекта генерацией ПД, в частоте и временном распределении которых кодируется вся зрительная инфа.
Проводниковый отдел вне сетчатки анатомичнески представлен вначале зрительными нервами, затем после частичного перекреста их волокон – зрительными трактами, а после преключения на нейронах латеральных коленчатых тел – зрительной лучистостью. От нейронов латеральных коленчатых инфа поступает в кору большого мозга. Часть зрительных путей проводит сигналы от сетчатки к РФ, ядрам гипоталамуса для управления циркадными биоритмами (сон/бодрствование), регуляции функций эндокринной и вегетативной НС; к нейронам претекальной зоны и верхних холмиков четверохолмия – для регуляции диаметра зрачка и аккомодации зрения через ядра и волокна вегетативной НС; для регуляции движений глаз через стволовые ядра и волокна 3, 4, 6 пар черепных нервов; к нейронам ядер вестибулярной системы и мозжечка для организации компенсаторных движений глаз при изменениях положения головы и тела в пространстве.
Корковый отдел системы зрения. Первичная сенсорная зона зрительной системы (стриарная кора) локализуется в затылочной области. В ней представлена ретинотопическая карта зрительных полей, играющая главную роль в формировании зрительных образов. Повреждение затылочных долей ведет к слепоте. Вторичная сенсорная кора расположена в экстрастриарной части (за пределами) первичной зрительной коре. Зрительная кора на каждой стороне мозга получает сигналы от ипсилатеральных (на той же стороне) половин сетчатки обоих глаз и коллатеральных половин полей зрения. В коре осуществляется слияние изображений от сетчатки обоих глаз в единое целое. Функциями зрительной коры являются обнаружение зрительного стимула, определение его формы, локализации в пространстве, контраста, размеров, цвета, направления движения и формирование зрительного образа. Восприятие других параметров осуществляется при участии экстрастриальных областейзрительной коры, теменной, лобной и др отделов коры большого мозга.
Слуховой анализатор: назначение основных структурных элементов, воспринимаемые характеристики звука, механизм восприятия различной высоты и силы звука, микрофонный эффект улитки, определение местоположения источника звука (опыт).
Характеристики звуковых волн. Различают высоту, силу и тембр звука.
Высота (частота звука) характеризуется частотой звуковых колебаний, чем она чаще, тем высота выше. Звуки могут различаться по высоте тона и диапазону частот.
Сила звука определяется давлением на барабанные перепонки.
Тембр звукового сигнала зависит от спектра, т.е. состава дополнительных частот, которые сопровождают основной тон.
Периферический отдел системы слуха – ухо.
Наружное ухо (ушная раковина и наружный слуховой проход) улавливает звуковые волны, обеспечивает концентрацию их и направляет на барабанную перепонку. Охраняют барабанную перепонку от механических и температурных воздействий внешней среды.
Среднее ухо представляет собой полость, наполненную воздухом, в которой находятся слуховые косточки, от нее идет евстахиева труба. От наружного слухового прохода оно отделено мембраной (барабанной перепонкой). Рукоятка молоточка вплетена в барабанную перепонку, другой его конец образует суставную поверхность с наковальней, которая сочленена со стремечком. Стремечко прилегает к мембране овального окна. Усиление звука в среднем ухе происходит с большей по сравнению с овальным окном площади барабанной перепонки, за счет ушной раковины и длины рычагов слуховых косточек.
Интенсивность звукового давления может быть понижена за счет защитного рефлекторного механизма аккомодации, который выражается в напряжении мышцы, которая оттягивает стремечко от овального окна, и мышцы, напрягающей барабанную перепонку. Рефлекс замыкается на уровне стволовых отделов мозга.
В барабанной полости поддерживается давление, равное атмосферному: во время глотания или зевания открывается отверстие евстахиевой трубы, которая соединяет полость среднего уха с носоглоткой.
Внутренней ухо – система сообщающихся заполненных жидкостью каналов и полостей в лабиринте височной кости. Здесь расположены улитка и вестибулярный аппарат.
Улитка – спирально закрученный костный канал, имеющий 2,5 завитка и разделенный базальной мембраной и мембраной Рейснера на три лестницы, начинается от овального окна. Барабанная лестница начинается круглым окном, через геликотрему (отверстие в верхушке) соединяется с вестибулярной лестницей, оба канала заполнены перилимфой. Средняя лестница заполнена эндолимфой. Внутри среднего канала расположен кортиев орган с рецепторными клетками. , преобразующими механические колебания в РП. Звуковые колебания передаются со стремечка на овальное окно, а оттуда сообщаются перилимфе. Колебания перилимфы распространяется на базальную мембрану.
Слуховые рецепторы (волосковые клетки кортиева органа) – вторичные механорецепторы. Они представлены внутренними и наружными клетками. На каждой клетке расположено около 100 волосков. Стереоцилии рецепторных клеток пронизывают покровную мембрану кортиева органа.один ее край связан с базальной мембраной, другой свободно свисает. Колебания базальной мембраны с волосковыми клетками сопровождаются касанием стереоцилий покровной мембраны, деформацией волосков, что ведет к развитию РП.
РП формируется следующим образом. В результате деформации стереоцилий на их верхушках открываются механоуправляемые каналы. Ионы К, содержание которых в эндолимфе больше, чем в рецепторной клетке, входят в клетку согласно электрическому и концентрационному градиентам. РП обеспечивает высвобождение медиатора в базальной части волосковых клеток, синаптически связанных с дендритами нейронов спинального ганглия. Глутамат действует на постсинаптическую мембрану окончания дендрита спирального ганглия и иницинирует формирование на ней генераторного потенциала, обеспечивающего возникновение ПД в окончании дендрита. При действии РП и ГП может быть зарегистрирован микрофонный эффект улитки.
Происхождение микрофонных потенциалов является следствием взаимодействия РП волосковых клеток, ГП и, по – видимому, ПД нервных окончаний слухового нерва, образующих синапсы с рецепторами.
Проводниковый отдел системы слуха. Первый нейрон биполярный. Он расположен в спиральном ганглии улитки; его дендрит синаптически связан с волосковыми клетками кортиева органа. Под действием медиатора в постсинаптической мембране (окончание дендрита первого нейрона) развивается ГП, который обеспечивает возникновение ПД в первом перехвате Ранвье этих окончаний. Аксоны биполярных клеток спирального ганглия формируют улитковую часть нерва и вместе с преддверной частью формируют преддверно – улитковый нерв, направляющийся через внутренний слуховой проход к вторым нейронам.
Второй нейрон расположен в ядрах кохлеарного комплекса продолговатого мозга. Их аксоны после частичного перекреста подходят к медиальному коленчатому телу метаталамуса, где образуют синаптические связи с третьими нейронами. Возбуждение от третьих нейронов идет в кору большого мозга к четвертому нейрону пути слуховой чувствительности. Часть аксонов второго нейрона транзитом проходят медиальное коленчатое тело и направляются к нижним буграм четверохолмия, образуя синапсы с их нейронами. Совокупность этих нейронов образует центры, нисходящие пути от которых направляются к мотонейронам спинного мозга.
Корковый отдел системы слуха находится в верхней части височной доли больших полушарий. Различают первичную и вторичную слуховую кору. Нервные импульсы от медиального коленчатого тела направляются в основном в первичную кору, а вторичная может активироваться как афферентными влияниями, идущими от таламуса, так и импульсами, которые формируются в первичной зоне.
Механизмы оценки характера звука. Различение высоты звуков. Характер и локализация колебаний базальной мембраны меняется в зависимости от частоты звука. Так, звуки низкой частоты приводят в колебательные движения всю мембрану, но особенно сильно в области близкой к геликотреме – верхнему отделу улитки, где находятся более длинные волокна основной мембраны.
При действии тонов до 400 Гц осуществляется пространственно - частотное кодирование. Пространственное заключается в том, что инфа в виде нервных импульсов передается по опред волокнам слухового нерва. Частотное выражается тем, что частота афферентных импульсов в слуховом нерве повторяет частоту звуковых колебаний.
С увеличение частоты максимальная амплитуда колеьбаний основной мембраны смещается к нижней части улитки, где расположены более короткие волокна. При этом частота возникающих в слуховом нерве ПД не возрастает. Первичная слуховая кора топографически поделена на области, которые отвечают за восприятие звуков различной высоты: низкочастотные звуки воспринимаются передними отделами височной коры. Нейроны задних отделов слуховой коры воспринимают высокочастотные колебания.
Механизмы различения силы звуковых колебаний:
С увеличением силы звука увеличивается амплитуда колебаний базальной мембраны , что ведет к возрастанию амплитуды РП и частоты ПД, распространяющихся по слуховому нерву в корковый отдел системы слуха.
Чем звук сильнее, тем больше площадь колеблющейся базальной мембраны и соответственно большее число волосковых клеток одновременно вовлекается в колебательный процесс; увеличение рецептивного поле приводит к пространственной суммации в ЦНС.
В возбуждение вовлекаются менее возбудимые и более глубоко расположенные в кортиевом органе рецепторы (внутренние клетки).
Определение локализации источника звука возможно с помощью биноурального слуха (способность слышать одновременно двумя ушами). Он обеспечивает сравнение характеристик звуковых колебаний, полученных каждым ухом. Звуковая волна раньше поступает к уху, на стороне которого локализуется источник звука: оценивается разность во времени прихода одинаковых фаз звуковой волны к обоим ушам.
Вестибулярный анализатор: роль основных структурных элементов в оценке положения тела в пространстве и при его перемещении, схема рефлекторной дуги, отражающая роль вестибулярного аппарата в регуляции мышечного тонуса.
Вестибулярной системе принадлежит важная роль в пространственной ориентации человека поддержании, его позы и регуляции движений.
Вестибулярная система обеспечивает восприятие инфы о прямолинейных и вращательных ускорениях движения тела и изменениях положения головы в пространстве или всего тела, а также о действии земного притяжения.
Периферический (рецепторный отдел) представлен двумя типами морфологически различных рецепторных волосковых клеток вестибулярного аппарата, расположенного вместе с улиткой в лабиринте височной кости. Вестибулярный орган состоит из преддверия и трех полукружных каналов. Преддверия включает два мешочка: сферический (саккулюс) и аллиптический, или маточку (утрикулюс). Полукружные каналы расположены в трех взаимно – перпендикулярных плоскостях. Они открываются своими устьями в преддверие. Один из концов каждого канала расширен в виде ампулы. Все эти структуры образуют перепончатый лабиринт, заполненный эндолимфой. Между перепончатым и костным лабиринтов находится перилимфа. В мешочках преддверия имеются небольшие возвышения, называемые пятнами (макулами), а в ампулах полукружных каналов гребешками (кристами). Здесь находятся рецепторные эпителиальные клетки, имеющие на свободной поверхности тонкие, многочисленные волоски (стереоцилии) и один более толстый длинный волосок (киноцилию).
Рецепторные клетки преддверия покрыты отолитовой мембраной, а находящиеся в ампулах листовой мембраной. В эти мембраны погружены волоски рецепторных клеток. Возбуждение волосковых клеток происходит при изгибании стереоцилий в сторону киноцилий и открытия механочувствительных калиевых каналов. Калий поступает в цитоплазму и деполяризует мембрану. Возникающий РП вызывает выделение из волосковых клеток АХ, который действует на холинорецепторы постсинаптической мембраны окончания дендрита нервного волокна первого нейрона вестибулярной системы. На постсинаптической мембране возникает ГП, электрическое поле которого вызывает ПД в афферентном волокне вестибулярного нерва. При смещении стереоцилий в сторону противоположную от киноцилий происходит закрытие ионных каналов, гиперполяризация мембраны и понижение активности сенсорных волокон вестибулярного нерва.
Адекватным раздражителем для рецепторных клеток преддверия являются линейные ускорения и наклоны головы или всего тела, приводящие к скольжению отолитовых мембран и изгибанию волосков. Для рецепторных клеток ампул полукружных каналов адекватным стимулом являются угловые ускорения в разных плоскостях при поворотах головы или вращениях тела.
Проводниковый отдел представлен как афферентными (сенсорными), так и эфферентными (меняющими функциональную мобильность волосковых клеток) волокнами. Аксоны биполярных нейронов (первый нейрон вестибулярного пути) вестибулярных ганглиев направляются к вестибулярным ядрам (Бехтерева, Роллера, Швальбе и Дейтерса) продолговатого мозга. Нейроны (второй нейрон) этих ядер получают дополнительную инфу от рецепторов мышц и суставов, сетчатки, мышц глаза, мозжечка и сами посылают сигналы ко многим отделам ЦНС: спинногму мозгу, глазодвигательным ядрам, вестибулярным ядрам противоположной стороны мозга, мозжечку, РФ, гипоталамусу, таламусу, в постлатеральном ядре которого расположены третьи нейроны, аксоны которых образуют прямой путь в кору БМ.
Корковый отдел (первая вестибулярная зона) представлен нейронами (четвертые нейроны) рассеянными в коре теменной и височной долей – в постцентральной извилине, верхней височной извилине, верхней теменной дольке. Вторая вестибулярная зона в премоторной коре, в нее поступает инфа по непрямому вестибулоцеребелоталамическому пути.
Типы вестибулярных реакций.
Соматические реакции обеспечивают поддержание тонуса мышц, координацию движений, нужное положение тела в пространстве и соответствующие глазодвигательные реакции. К ним относят многие врожденные спинальные и стволовые рефлексы – статические, статокинетические и кинетические. Вертикальная поза и походка определяются преимущественно отолитовым аппаратом, а глазодвигательные реакции вызываются с рецепторов полукружных каналов. Вестибулярный нистагм (статокинетический рефлекс) – особая форма глазодвигательных реакций, необходимая для удержания объекта наблюдения в поле зрения при поворотах головы или вращения тела.
Сенсорные (корковые реакции) возникают при определенной интенсивности раздражений и выражаются чувством головокружения, разнообразными пространственными иллюзиями собственного положения или окружающих предметов относительно друг друга.
Вестибулярно – вегетативные реакции обеспечивают необходимый в данный момент времени уровень обменных процессов в мышечной системе.
Кинетоз – болезнь передвижения (морская, воздушная).
Тактильный и температурный анализаторы: структурно-функциональная их характеристика, значение.
Система температурной чувствительности участвует в поддержании гомеостазиса с помощью изменения интенсивности теплопродукции и теплоотдачи, а также поведенческих приспособительных реакций, если температура внешней среды становится некомфортной.
Периферический отдел представлен холодовыми и тепловыми терморецепторами кожи: тепловые – тельца Руффини, холодовые - колбы Краузе. Рецепторы холода расположены в эпителии или непосредственно под ним, рецепторы тепла глубже – в сосочковом слое дермы. Терморецепторы расположены также в слизистой полости рта, холодовые преобладают в передних отделах слизистой, тепловые в задних.
Высокой чувствительностью к температурным раздражениям обладают кончик языка и красная каемка губ, т.к. при приеме пищи в первую очередь раздражаются именно эти области.
Терморецепторы внутренних органов воспринимают изменения температуры внутренней среды организма, что определяет их ведущую роль в поддержании постоянства температуры внутренней среды.
Проводниковый отдел представлен тонкими немиелинизированными волокнамитипа дельта (заканчиваются холодовыми рецепторами) и безмиелиновыми волокнами типа С (их окончания – рецепторы тела). Чувствительные спинальные ганглии и ганглии черепных нервов, где расположен первый нейрон, являются источником нервов, иннервирующих различные участки кожи тела, слизистой оболочки полости рта. Клетки задних рогов спинного мозга и ядер черепных нервов ствола мозга представляют второй нейрон. Нервные волокна, отходящие от вторых нейронов системы внешней температуры в составе латерального спиноталамического тракта, направляются к зрительному бугру, где образуют синаптические контакты с третьими нейрономи. Отсюда инфа о температуре внешней среды поступает в кору (четвертый нейрон).
Корковый отдел локализуется в области задней центральной извилины коры головного мозга и соответствует локализации тактильной чувствительности.
Тактильная система обеспечивает формирование ощущений прикосновения, давления, вибрации и щекотки.
Периферический отдел представлен семью видами рецепторов, различающихся строением и локализацией. Рецепторы, воспринимающие давление: диски Меркеля, расположенные небольшими группами в глубоких слоях кожи и слизистых оболочках; тельца Руффини в глубоких слоях эпидермиса и глубоких слоях дермы; концевые колбы Краузе под эпидермисом, в соединительнотканной основе слизистых оболочек и среди мышечных волокон языка; свободно лежащие неинкапсуированные нервные окончания в поверхностных слоях кожи. Рецепторы прикосновения: тельца Мейснера в базальном и сосочковом слоях кожи, на поверхности кожи лишенной волос, слизистых оболочках; рецепторы волосяного фолликула. Восприятие вибрации: тельца Пачини в слизистой, коже, не покрытой волосами, в жировой ткани подкожных слоев, в суставных сумках и сухожилиях.
Проводниковый отдел начинается от механорецепторов А – волокнами. Первый нейрон локализуется в спинальных ганглиях или ганглиях черепных нервов. Инфа идет в спинной мозг и ствол мозга. В заднем роге спинного мозга аксоны нейронов спинальных ганглиев в составе задних канатиков спинного мозга восходят к продолговатому мозгу, где в ядрах заднего столба происходит первое переключение (второй нейрон). От кожи головы и слизистой рта импульсы идут по тройнично – таламическому тракту: первый нейрон – в тройничном ганглии, второй нейрон – в главном ядре средней части моста. Далее через медиальную петлю проводящий путь тактильной системы следует к коллатеральным вентробазальным ядрам зрительного бугра (третий нейрон, второе переключение). Нейроны таламических ядер посылают свои аксоны в корковый отдел системы (четвертый нейрон, третье переключение).
Корковый отдел находится в 1 и 2 зонах постцентральной извилины (соматосенсорная область коры). От проекционных зон коры тактильная инфа поступает во фронтальные и задние ассоциативные зоны коры.
Ноцицепция: биологическое значение боли, проекционные и отраженные боли, ноцицептивная и антиноцицептивная системы.
Боль – неприятные в виде страданий ощущения, возникающие в результате сверхсильного раздражения, повреждения тканей, возникновения патологического процесса и кислородного голодания тканей.
Физиологическая роль боли заключается в следующем:
Выполняет роль сигнала об угрозе или повреждении тканей организма и предупреждает их.
Является фактором мобилизации защитно-приспособительных реакций при повреждении его органов и тканей
Имеет познавательную функцию: через боль человек, начиная с раннего детства, учится избегать возможные опасности внешней среды.
Эмоциональный компонент боли выполняет функцию подкрепления при образовании условных рефлексов даже при однократном сочетании условного и безусловного раздражителей.
Болевая система (ноцицептивная).
В болевой реакции выделяют следующие компоненты: 1. Сенсорный, отражающий возникновение ощущения боли; 2. Эмоциональный в виде страдания, тягостных переживании, формирующихся на основе иррадиации ретикулярных возбуждений к структурам лимбической, теменной и лобной коры; 3. Вегетативный, проявляющийся в изменении практически всех вегетативных эффекторов; 4. Двигательный, выступающий в виде рефлекса избегания, локального или генерализационного напряжения определенных групп мышц, в других формах двигательной активности организма.
Типы болей.
По месту возникновения разделяют на соматическую и висцеральную. Боль, возникающая в коже – соматическая поверхностная боль. Боль, возникающая при поражении суставных связок, мышц, суставов, надкостницы, костей – соматическая глубокая боль. Причины возникновения висцеральной боли: 1. Ишемия тканей, приводящая к накоплению метаболитов и нарушению тканевого дыхания; 2. Выход содержимого полых органов при повреждении целостности стенок; 3. Сильные тонические сокращения или растяжения мышц желудка, желчного пузыря, кишечника, мочевого пузыря, мочеточников.
По продолжительности острая, подострая и хроническая. Острая – внезапно возникающая боль. Хроническая – длительная, долго сохраняющаяся или регулярно повторяющаяся боль.
По отношению к причинам возникновения различают физическую и психогенную.
Проекционная боль – боль в зоне иннервации периферического нерва, возникающая при раздражении его ствола, напр. патологическим процессом.
Отраженными болями называются болевые ощущения не во внутренних органах, от которых поступают болевые сигналы, а в определенных частях кожной поверхности (зоны Захарьина-Геда). Так, при стенокардии кроме болей в области сердца ощущается боль в левой руке и лопатке. Возникновение этих болей связано с тем, нейроны, проводящие болевые импульсы от рецепторов пораженного органа и рецепторов соответствующего участка кожи, конвергируют на одном и том же нейроне спиноталамического пути.
Периферический отдел системы боли. Ноцирецепторы представляют собой свободные, неинкапсулированные окончания нервных волокон.
Большинство ноцирецепторов полисенсорны: реагируют на механические, термические и химические раздражители.
В случае механического воздействия на мембрану ноцирецептора РП возникает при ее деформации, активирующей поступление натрия внутрь рецептора. В случае термического воздействия формирование РП протекает, очевидно, с участием медиатора, выделяющегося из терминалей сенсорного нейрона под влиянием температурного воздействия. Медиатор вступает в реакцию с хеморецептором, запускает генерацию РП.
При воспалительных реакциях ощущение боли возникает в результате активации различных видов хемононицепторов алгогенами – хим в – вами, действующими на хемоноцицептор в составе своеобразного химического конвейера. В него входят тканевые алгогены – гистамин, серотонин, АХ, простагландины, появляющиеся в межклеточном пространстве в результате угнетения клеточного дыхания, деструкции тканевых клеток, нервных волокон, а также плазменные алгогены – каллидин, брадиеинин, появляющиеся в результате выхода в межклеточное пространство компонентов плазмы крови. Большинство алгогенов оказывают прямое активирующее влияние на мембрану хемоноцирецептора, некоторые (простагландины) обеспечивают модулирующее влияние, резко повышая чувствительность хемоноцирецептора к активирующим его агентам.
Проводниковый отдел болевой системы представлен тонкими миелинизированными волокнами типа а и тонкими немиелинизированными волокнами типа С.
Болевая импулльсация по волокнам А в составе задних корешков поступает в задние рога спинного мозга, в которых через глутаматергические синапсы происходит передача возбуждения с нейронов первого порядка на нейроны второго порядка. Аксоны нейронов второго порядка переходят в противоположную сторону, направляясь в ствол мозга в составе переднего спиноталамического тракта. В стволе мозга меньшая часть волокон вступает в контакт с нейронами ядер РФ, большая вступает в таламус, где происходит передача возбуждения на нейроны третьего порядка в составе специфического комплекса таламических ядер.
Болевая импульсация по волокнам типа С поступает в задние рога в составе задних корешков. Большая часть переключается здесь на нейроны второго порядка, аксоны которых переходят на противоположную сторону, направляясь в ствол мозга в составе латерального спиноталамичечского тракта. Волокна латерального спиноталамического тракта на уровне ствола мозга вступают в контакт с различенными нейронными структурами. Среди них РФ продолговатого мозга, моста, среднего мозга, нейроны голубого пятна. Многократно переключаясь на ретикулярных нейронах с короткими аксонами, возбуждения направляются в неспецифические ядра таламуса. Частично возбуждения направляются в специфические ядра, в структуры гипоталамуса и др подкорковых образований мозга. Часть волокон латерального спиноталамического пути вступают в неспецифические ядра таламуса, проходя ствол мозга без переключений.
Корковый отдел. Аксоны нейронов специфических ядер таламуса предают возбуждение в первичное проекционное соматосенсорное поле SI, расположенное в области постцентральной извилины, и во вторичное проекционное соматосенсорное поле SII, расположенное в глубине сильвиевой борозды. Аксоны нейронов неспецифических ядер передают возбуждение ко всем участкам коры, а также к структурам лимбической системы.
Поле SI обеспечивает восприятие быстрой боли, идентификацию места ее возникновения на теле. Точная идентификация участка кожной поверхности, на которую действует раздражитель, требует подключения кожных тактильных рецепторов. Полю SI отводится ведущая роль в экстренном включении моторной защитной реакции организма в ответ на действие болевого раздражителя. Эта реакция обеспечивается прямыми синаптическими связями нейронов поля SI с нейронами момторной коры.
Нейроны поля SII имеют двусторонние связи с ядрами таламуса, что позволяет ему избирательно фильтровать проходящие через таламус возбуждения, выделяя из них возбуждения болевого характера. После этого нейроны поля SII вовлекаются в многостадийные процессы аналитико – синтетической деятельности мозга, связанные с из извлечением из памяти программ требуемых поведенческих актов, обеспечением их реализации на эффекторах, проведением оценки качества достигнутого в последовательности поведенческих актов полезного результата.
Активность полей SI и SII связана с активностью других функциональных полей коры, в том числе и лобной доли.
Обезболивающая система.
Ощущения боли впервые обнаруживаются при некоторой минимальной силе раздражителя, получившей название болевого порога. Сила раздражителя, при которой испытуемый просил прекратить действие, получила название порога болеустойчивости. Гипералгезия- повышение болевой чувствительности, гипоалгезия – понижение, аналгезия – нечувствительность к боли.
Уровни обезболивающей системы.
Стволовой уровень включает центральное серое околоводопроводное вещество (ЦСОВ) и ядра шва (ЯШ), образующие единый функциональный комплекс. Изолированная электрическая стимуляция ЦСОВ – ЯШ блокирует проведение болевой импульсации на уровне релейных нейронов ядер задних рогов спинного мозга, а также релейных нейронов сенсорных ядер тройничного нерва.
Специфические влияния ЦСОВ – ЯШ на соответствующие нейроны реализуются нисходящими аксонами, большинство из которых в своих окончаниях выделяют серотонин. Он тормозит активность релейных нейронов, воспринимающих болевую импульсацию с периферии, но вызывает возбуждение энкефалинергических нейронов. Возбуждение энкефалинергических нейронов обеспечивает включение дополнительных механизмов пост и пресинаптического торможения релейных нейронов.
Также в состав антиноцицептивных механизмов стволового уровня входят парагигантоклеточное ядро РФ ствола мозга (ПГЯ) и голубое пятно (ГП). Наибольшая плотность аксонов нейронов ПГЯ и ПГ наблюдается в местах расположения сенсорных ядер тройничного нерва. Стимуляция ПГЯ и ПГ подавляет болевые ответы нейронов этих ядер. Механизмы подавления пост и пресинаптические за счет энкефалинергических и норадреналинергических влияний на рецепторы соматической мембраны релейных нейронов, а также на рецепторы мембран пресинаптических С – волокон.
Активация стволовых механизмов антиноцицептивной системы происходит за счет нарастания интенсивности восходящего болевого потока, поступающего по спиноталамическим и тригеминоталамическим трактам в таламус. По коллатералям волокон этих трактов импульсация возбуждает нейроны гигантоклеточного ядра (ГЯ) и ПГЯ. Последние возбуждают практически весь комплекс стволовых антиноцицептивных механизмов.
Гипоталамический уровень. Раздражение гипоталамической зоны положительного подкрепления вызывает гипоалгезию – частично за счет бета – эндорфинергических и энкефалинергических влияний, усиливающих активность антиноцицептивных механизмов стволового уровня, частично за счет бета – эндорфинергических влияний, угнетающих прохождение болевой импульсации в неспецифических ядрах таламуса.
Раздражение гипоталамической зоны отрицательного подкрепления сопровождается гипоалгезией. Пусковая роль в возникновении гипоалгезии отводится норадреналинергических и дофаминергических гипоталамическим нейронам, контролирующим антиноциицептивные механизмы стволового уровня, предположительно связанные с активностью ПГЯ и ГП. При активации норадреналинергических и дофаминергических нейронов гипоталамической зоны отрицательного подкрепления сопровождается параллельной активацией клеток передней доли гипофиза. В результате в кровь начинает поступать бета – эндорфин, вторично оказывающий мощное угнетающее влияние на нейроны всех каналов передачи болевой импульсации в ЦНС.
Корковый уровень. Нисходящие волокна коркового контроля болевой чувствительности, направляющиеся к задним рогам спинного мозга и сенсорным ядрам тройничного нерва, берут начало в различных областях моторной, орбитальной коры, в полях SI, SII. Кортикофугальные влияния на нейроны могут быть возбуждающими, однако преобладают пре и постсинаптические тормозные влияния. Помимо прямых влияний на нейроны спинного мозга и тройничного нерва поле SII контролирует их влияние опосредовано, модулирую активность стволового комплекса антиноцицептивной системы.
Ведущее место в обеспечении кортикогипоталамических влияний отводится лобной коре. Характер ее влияния на гипоталамус модулирующий: в одних случаях активность ядер, участвующих в обработке болевой импульсации усиливается, в других снижается.
Субъективное восприятие боли зависит от баланса активностей ноцицептивной и антиноцицептивной систем. Повышение функциональной активностиноцицептивной системы или снижение активности антиноцицептивной системы приводит к увеличению болевой чувствительности.
Обонятельный и вкусовой анализаторы: их значение; периферические, проводниковые и центральные отделы этих анализаторов.
Система вкуса позволяет формировать вкусовые ощущения. Чувство вкуса многокомпонентно и связано с раздражением механо, термо, хемо и даже болевых рецепторов слизистой оболочки полости рта и глотки. Главным назначением системы вкуса является определение пригодности и ценности пищи, а также в формировании аппетита.
Ведущую роль в формировании вкусовых ощущений выполняет хеморецепция. Известно 13 видов хеморецепторных клеток, чувствительных к разным веществам, таким кА к натрий, калий, хлор, глутамат, ионы водорода и т.д. активация этих рецепторов вызывает 4 типа вкусовых ощущений: сладкое, соленое, горькое, кислое.
Порог чувствительности для горьких веществ самый низкий, т.к., как правило, горьким вкусом обладают несъедобные или ядовитые вещества.
Периферический отдел. Вкусовые рецепторы являются вторичными. Они представляют собой клетки с микроворсинками, обладают спонтанной активностью, совместно с опорными клетками и нервными окончаниями формируют вкусовые почки. Узкая часть представлена микроворсинками вкусовых клеток, на которых расположены рецепторы. Микроворсинки контактируют с содержанием ротоглотки через небольшие отверстия в слизистой – вкусовые поры. Вкусовые поры локализуются на трех типах сосочков: окруженных валом (в области корня языка), грибовидных (на спинке) и листовидных (на складках боковых поверхностей). Кончик языка и передняя 1/3 чувствительны к сладкому, корень к горькому, бок поверхности к кислому и соленому. Взаимодействие рецепторов ворсинок с химическими веществами ведет к открытию натриевых каналов и деполяризации, вследствие чего формируется РП, высвобождается медиатор, под влиянием которого в чувствительном нервном окончании (нейроны дендритов ганглиев черепных нервов) возникает ГП. Электрическое поле ГП вызывает ПД.
Проводниковый отдел начинается внутри вкусовой почки, где находятся синапсы рецепторных клеток и афферентных нервных волокон. В афферентных волокнах имеется фоновая импульсация вкуса (без действия раздражителя).
Вкусовые почки, расположенные в передних 2\3 языка, иннервируются волокнами барабанной струны, входящей в состав лицевого нерва и язычного нерва; почки задней 1\3 иннервируются, твердого и мягкого неба, миндалин получают волокна от языкоглоточного нерва; вкусовые почки надгортанника, гортани глотки иннервируются верхнегортанным нервом (порция блуждающего нерва).
Афферентные волокна этих нервов являются дендритами биполярных нейронов (первый нейрон), расположенные в соответствующих чувствительных ганглиях черепных нервов. Аксоны этих нейронов входят в состав одиночного пучка продолговатого мозга, в ядрах которого локализуются вторые нейроны. Аксоны вторых нейронов в составе медиальной петли подходят к таламусу, где локализуются третьи нейроны, посылающие свои аксоны к коре, где локализуются четвертые нейроны.
Корковый отдел находится в нижней части постцентральной извилины (соматосенсорная зона коры) в области представительства языка.
Система обоняния позволяет определить нахождение в воздухе пахучих веществ. Она способствует ориентации организма в окружающей среде и процессупознания внешнего мира, помогает избежать опасностей благодаря способности различать вредные для организма вещества, оказывает влияние на деятельность ЖКТ.
Периферическим отделом являются дендриты биполярных обонятельных нейронов, расположенных в слизистой оболочке носовых ходов, которая в этой части называется обонятельной мембраной. Терминали дендритов обонятельных нейронов представлены 6 – 12 ресничками, в мембранах которых локализуются специфические для определенных одорантов белки – рецепторы. Активация мембранных рецепторов ресничек приводит к синтезу цАМФ, при участии которого открываются натриевые каналы и в дендрите обонятельного нейрона формируется РП. Электрическое поле РП обеспечивает деполяризацию аксонного холмика и тела обонятельного нейрона, которая, достигнув КП, ведет к возникновению ПД.
Проводниковый отдел: аксоны обонятельных нейронов (первые нейроны), пронизывающие пластинку решетчатой кости, взаимодействуют с крупными митральными клетками (вторые нейроны) в области, называемой обонятельной луковицей, расположенной в коре вентральной поверхности лобной доли. Аксоны митральных клеток обонятельных луковиц образуют обонятельный тракт, инфа по которому доставляется к корковому отделу.
Корковый отдел включает прегрушевидную область, обонятельную извилину гиппокамп. Большое количество нервных волокон из перечисленных образований направляется снова к обонятельным луковицам. Они заканчиваются на гранулярных клетках гломерул. Гранулярные клетки образуют тормозные синапсы с митральными клетками.
Двигательный (кинестетический) анализатор: общая характеристика, роль в регуляции мышечного тонуса и осуществлении шагательного рефлекса (схема, уровень спинного мозга).
Кинестетический анализатор (проприоцептивная система) обеспечивает возникновение ощущений о положении тела и его частей в пространстве в результате действия силы гравитации и сокращения или расслабления скелетных мышц, участвует в координации мышечной деятельности.
Периферический отдел представлен первично чувствующими рецепторами мышц, сухожилий, суставных сумок. Рецепторы мышц расположены в мышечных веретенах параллельно рабочим мышечным волокнам. Выделяют два типа мышечных веретен: ядерно – сумчатые и ядерно – цепочечные, и два типа мышечных рецепторов: быстроадаптирующиеся (реагируют на скорость изменения длины мышц) и медленно адаптирующиеся (оценивают длину мышцы). Сухожильные рецепторы (рецепторы Гольджи) подключены относительно экстрафузальных мышечных волокон последовательно и служат для оценки напряжения мышцы и регуляции силы ее сокращений. Рецепторы суставных сумок реагируют на положение суставов и изменение суставного угла.
Проводниковый отдел представлен тремя основными путями: лемнисковым, спиномозжечковыми и проприоспинальными. Лемнисковый путь включает чувствительные нейроны спинномозговых узлов (первый нейрон); из аксоны в составе задних канатиков спинного мозга достигают нейронов ядер Голля и Бурдаха (второй нейрон), аксоны которых в составе медиальной петли (лемниск) после перекреста направляются в специфические ядра таламуса (третий нейрон). Аксоны таламических нейронов достигают нейронов коры (четвертый нейрон)
Корковый отдел представлен нейронами первой и второй симатосенсорных областей постцентральной извилины, которые имеют соматотопическую организацию.
Внутренние (висцеральные) анализаторы: классификация, роль в поддержании гомеостазиса, в регуляции функции внутренних органов.
Висцеральные сенсорные системы выполняют функцию получения, анализа и синтеза инфы о состоянии внутренней среды организма, тем самым они участвуют в регуляции работы внутренних органов. Выделяют следующие системы внутренней среды: химизма, температуры, осмотического давления, давления в кровеносных сосудах, давления во внутренних полых органах при их наполнении.
Периферический отдел представлен соответствующими рецепторами (механо, термо, хемо и осмо), расположенными в различных органах, сосудах, слизистых оболочках и ЦНС. Все они являются первичными. Адекватным стимулом для них являются различные химические вещества.
Проводниковый отдел: афферентная импульсация от внутренних органов поступает в ЦНС в основном в составе вегетативных нервов, частично в смешанных, в состав которых входят и соматические, и вегетативные волокна. Первые нейроны локализуются в соответствующих чувствительных ганглиях, вторые в спинном или продолговатом мозге. Их аксоны идут к заднемедиальному ядру таламуса (третий нейрон).
Корковый отдел находится в зонах С1 и С2 соматосенсорной области коры (четвертый нейрон) и в орбитальной области коры большого мозга. Поступление потока висцеральной импульсации к нейронам коры обеспечивает возможность выработки многочисленных вегетативных условных рефлексов.
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ. АДАПТАЦИЯ И ЗАЩИТНЫЕ СИСТЕМЫ ОРГАНИЗМА
Высшая нервная деятельность (ВНД): понятие, особое значение учения о ВНД для клиники, развитие представления о ВНД (Гольц, И.М.Сеченов, И.П.Павлов, Э.А.Асратян, П.К.Анохин - схема “поведенческой“ функциональной системы. Методики изучения ВНД.
Основоположником учения о ВНД является И.П.Павлов, открывший условные рефлексы. Используя их как объективный метод исследования психической деятельности, он разработал основы учения о ВНД.
История изучения ВНД.
Экспериментальной предпосылкой для создания учения о ВНД были работы по декортикации (полной или частичной) подопытных животных с последующими наблюдениями за изменениями в поведении (Буйо, Флуранц, Гольц). Л. Гольц в течение 18 месяцев наблюдал за поведением собаки с удаленными большими полушариями. Животное потеряло все навыки, приобретенные в течение жизни, в результате чего не могло приспособиться к изменениям внутренней среды.
И. М. Сеченов высказал мысль о том, что сознание представляет собой лишь отражение реальной действительности. Познание окружающей среды возможно лишь с помощью органов чувств – первоначального звена всей психической деятельности. Первопричина всякого действия лежит вне его, наблюдая за поведением и формированием сознания ребенка, он показал, как врожденные рефлексы с возрастом усложняются, вступают в разнообразные связи друг с другом и определяют всю сложность человеческого поведения.
Условный рефлекс является объективным методом исследования ВНД мозга. Исследование условно – рефлекторного слюноотделения стало отправной точкой в создании Павловым учения о ВНД.
ВНД, по Павлову – условно – рефлекторная деятельность при участии ведущих отделов головного мозга ( у человека и животных больших полушарий и переднего мозга), обеспечивающих адекватные и наиболее совершенные отношения организма к вешнему миру, т.е. поведение. Вместе с тем обработка полученной информации в период бодрствования, а также во сне являются проявлением ВНД, хотя активное поведение при этом отсутствует. ВНД – те процессы, которые происходят в мозге и обеспечивают приспособление.
ВНД – совокупность нейрофизиологических процессов, обеспечивающих сознание, подсознательное усвоение инфы и обучение в онтогенезе всем видам деятельности, в том числе целенаправленному поведению организма в окружающей среде и обществе. Психическая деятельность – идеальная, субъективно осознаваемая деятельность организма, осуществляемая с помощью нейрофизиологических механизмов. Психическая деятельность осуществляется с помощью ВНД.
Существенный вклад в развитие учения о ВНД внесли ученики Павлова. Асратян открыл условные рефлексы с прямыми и обратными связями.
Велики заслуги Анохина, который разработал схему архитектоники поведенческого акта.
Методы исследования ВНД.
ВНД изучается с помощью выработки условных рефлексов и др видов научения , методами воздействия на ЦНС и исследования ее состояния.
Выработка различных условных рефлексов и др форм научения на фоне холодового выключения, разрушения и раздражения отдельных структур мозга позволяет выявить роль отдельных структур в замыкании временной связи.
Научение проводят при введении агонистов (АХ, адреналина, серотонина), антагонистов (блокаторов), а также на фоне действия молекул ДНК и РНК.
С помощью электронной микроскопии исследуют изменения в синаптических структурах и отростках нейронов после выработки условных рефлексов и при исследовании др видов научения.
Электроэнцефалография – регистрация суммарного электрического поля, создаваемого ПД, ВПСП и ТПСП, коры большого мозга с поверхности головы. ЭЭГ с коры - электрокортикограмма. Метод ЭЭГ позволяет в некоторой степени судить о локализации фокуса активности. Регистрация ЭЭГ проводится с помощью биполярных или униполярных электродов, накладываемых на кожу головы. На ЭЭГ можно выделить 4 основных ритма: альфа, бета, тета, дельта.
Микроэлектродным методом изучается активность одиночных нейронов ЦНС. С помощью микроэлектродов, вводимых внутрь клетки, можно измерять мембранные ПП, регистрировать постсинаптические потенциалы – возбуждающие и тормозные, а также ПД. Разновидностью микроэлектродного метода является метод микроинофореза, при котором используются многоканальные стеклянные микроэлектроды. Через один канал, заполненный электролитом, можно регистрировать электрическую активность нейрона; остальные каналы заполняют БАВ, которые апплицируют на работающий нейрон, пропуская растворы веществ через постоянный ток.
На базе метода магнитно – резонансной томографии (МРТ), которая применяется для структурной томографии (получение карты структур мозга на основе контрастов серого и белого веществ), появилась функциональная МРТ. Она основана на использовании парамагнитных свойств веществ, которые можно ввести в организм и которые не обладают магнитными свойствами, но приобретают их, войдя в магнитное поле. На практике ФМРТ использует парамагнитные свойства гемоглобина. ФМРТ позволяет выявлять участки мозга с активно работающими нейронами.
Классификация условных рефлексов и их характеристика. Причины более совершенного приспособления организма в окружающей среде с помощью условных рефлексов. Импринтинг: понятие, формы, отличительные особенности.
Среди условных рефлексов, которые вырабатываются на базе безусловных необходимо выделить три группы.
По биологическому значению выделяют пищевые, оборонительные, половые.
По виду рецепторов, с которыми вырабатываются условные рефлексы выделяют:
Экстероцептивные условные рефлексы формируют приспособительное поведение в окружающей среде
Интероцептивные участвуют в регуляции функций внутренних органов.
Проприоцептивные лежат в основе научения двигательным навыкам
По отделу НС и эффекторам различают условные соматические (двигательные) и вегетативные (сердечно –сосудистые, секреторные, выделительные).
В зависимости от условий выработки различают:
Натуральные условные рефлексы образуются на сигналы, являющиеся естественными признаками подкрепляющего раздражителя.
Искусственные (лабораторные) – условные рефлексы на любые сигнальные раздражители, которые в природе не имеют отношения к безусловному раздражителю. Эти рефлексы разделяют на три основные подгруппы.
По сложности различают: 1. Простые условные рефлексы, вырабатываемые на одиночные раздражители; 2. Комплексные условные рефлексы: один условный рефлекс на несколько сигналов, действующих одновременно или последовательно; 3. Цепные рефлексы – несколько условных рефлексов на цепь раздражителей, каждый из которых вызывает свой условный рефлекс – динамический стереотип, иллюстрирующий системность в деятельности коры большого мозга, аналитико – системную деятельность; 4. Инструментальные условные рефлексы: например, животное в эксперименте для получения пищи должно начать на педаль.
По соотношению времени действия условного и безусловного раздражителей различают наличные и следовые. Для выработки наличных условных рефлексов характерно совпадение действия условного и безусловного раздражителей, последний включается несколько позже. Следовые рефлексы вырабатывают в условиях, когда безусловный раздражитель подают через 2- 3 мин после выключения уловного.
По выработке условного рефлекса на базе другого условного рефлекса различают условные рефлексы второго, третьего и др порядков. Рефлексы первого порядка - рефлексы, выработанные на базе безусловных рефлексов. Рефлексы второго порядка вырабатываются на базе рефлексов первого порядка, при которых безусловный стимул отсутствует. Рефлекс третьего порядка на базу рефлекса второго порядка.
У человека вырабатываются условные рефлексы на слово.
Выделяют также положительные и отрицательные условные рефлексы (торможение условных рефлексов). Однако такое разделение некорректно. Условный рефлекс – выработанная в процессе онтогенеза реакция организма на раздражитель, ранее индифферентный для этой реакции и носит приспособительный характер, что не может быть отрицательным явлением.
Импринтинг – запечатлевание в памяти новорожденного окружающей действительности. Это особая форма неассоциативного научения.
Формы импринтинга.
Запечатление образов и объектов – родителей, братьев, сестер, вида пищи и др.
Усвоение поведенческих актов – имитационное поведение.
Реакция следования – автоматическое следование новорожденного за родителем.
Импринтинг имеет сходство с безусловным и условным рефлексами. Подобно безусловным рефлексам он сохраняется, как правило, на всю жизнь. Реакции импринтинга являются врожденными, но для их проявления нужны определенные условия.
Импринтинг отличается и от безусловных, и от условных рефлексов. Во – первых, он проявляется в определенные критические периоды. Имеется три основных критических периода: 1. Критический период для запечатления родителя – первые часы (максимум дни) жизни детеныша; 2. Импринтинг у матери по отношению к новорожденному; 3. Формирование полового поведения и предпочтения будущего полового партнера.
Во – вторых, запечатление происходит очень быстро и возможно с первого раза.
Биологическое значение импринтинга заключается в том, что он выделяет и позволяет усваивать ключевые моменты зоосоциального поведения, которые являются настолько важными, что их надо формировать быстро и надолго.
Научение: понятия, научение в фило- и онтогенезе, роль отдельных структур мозга, основные формы научения.
Научение - выработка в онтогенезе приспособительных форм поведения, в том числе и навыков физического труда. Выработка условного рефлекса тоже научение.
Условный рефлекс – выработанная в онтогенезе реакция организма на раздражитель, ранее индифферентный для этой реакции.
Рефлексы подчиняются закону силы – с увеличением силы стимула ответная реакция возрастает; включают афферентную, центральную и эфферентную части дуги; в биологическом значении являются приспособительными.
На один и тот же раздражитель можно вырабатывать различные условные рефлексы (условно – рефлекторное переключение).
Особое биологическое значение условных рефлексов в том, что они обеспечивают более тонкое приспособление организма. Во – первых, они возникают заблаговременно, готовя организм к будущей полезной поведенческой деятельности и помогая ему избежать вредных воздействий, тонко и эффективно адаптироваться к окружающей среде. Во – вторых, их набор неограничен; в – третьих, они изменчивы: формируются и возникают в определенной ситуации и угасают, если ситуация меняется.
Все виды научения подразделяют на две основные группы: неассоциативное (привыкание, сенситизация, импринтинг и подражание) и ассоциативное. Неассоциативное заключается в реагировании или не реагировании на раздражитель без использования сигнала, без связи (ассоциации), т.е. без совпадения с каким – то сигналом. Ассоциативное формируется при совпадении индифферентного раздражителя с деятельностью организма. При этом формируется связь (ассоциация) между сигналом и раздражителем (подкреплением). Также выделяют пассивное, оперантное, научение с помощью мышления, подсознательное.
Пассивное (реактивное) научение. Этот вид научения имеет место во всех случаях, когда организм приобретает навыки пассивно, т.е. не прилагая целенаправленных усилий. К нему относятся привыкание, сенситизация, условные рефлексы, импринтинг.
Привыкание (габитуация) – угасание ориентировочной реакции. Если раздражитель многократно повторяется и не имеет значение для организма, то организм прекращает на него реагировать.
Сенситизация – усиление реакции организма на повторяющийся стимул, который каждый раз вызывает неприятные ощущения. Научение выражается в соответствующих поведенческих реакциях типа стимул – ответ (после нескольких повторений действия раздражителя). В основе механизма этого вида лежит механизм синаптической потенциации.
Условные рефлексы (см. ранее)
Импринтинг (см. ранее).
Оперантное научение. В ходе этого научения организм добивается полезного результата с помощью активного поведения. Имеется три основных вида: метод проб и ошибок, инструментальный условный рефлекс, самораздражение или его избегание.
Метод проб и ошибок. Кошек помещали в проблемные камеры, которые открывались при определенных действиях кошки. Когда кошка выходила ей давали пищу. По мере повторения процедуры выхода из клетки (увеличении числа проб и ошибок) скорость выполнения задачи возрастала.
Инструментальный условный рефлекс – научение действию по сигналу (в первом методе без сигнала) с помощью вознаграждения. Крыса придействии света нажимает на рычаг, чтобы получить пищу.
Самораздражение структур мозга для получения удовольствия, положительных эмоций или избегание самораздражения при неприятных ощущениях, возникающих от раздражения.
Когнитивное научение (научение с помощью мышления). К нему относится научение путем наблюдения и правильное действие организма в ситуации, с которой он впервые встретился.
Научение путем наблюдения – подражательное и викарное. Викарное научение осуществляется посредством наблюдения, но при этом результат действия заранее оценивается, т.е. выполняется с определенной целью.
Научение в ситуации, впервые встретившейся организму. Например, экстраполяционные рефлексы, заключающиеся в способности животных определять направление движения предмета или другого животного, движущегося по прямой линии после исчезновения их из поля зрения.
Подсознательное научение. Внезапное правильное решение задачи с помощью подсознательной деятельности мозга.
Виды и механизмы памяти; мгновенная, кратковременная и долговременная память, и опыты их доказывающие. Механизм образования условных рефлексов, схема, ее иллюстрирующая. “Замыкание” временной связи - (Э.А.Асратян).
Память – способность организма приобретать, сохранять и формировать в сознании информацию и навыки. Различают врожденную – генетическую и приобретенную – иммунную и нейронную память.
Классификация памяти. По формам восприятия инфы различают слуховую и моторную; по уровням освоения – воспроизводящую и облегчающую; по длительности хранения информации - кратковременную и долговременную. Выделяют также ассоциативную и неассоциативную, процедурную и декларативную. Процедурная связана с усвоением навыков. Декларативная – хранение любой инфы (кроме навыков): формул, символов, содержание прочитанной книги.
Хранение инфы в долговременной памяти называется энграммой. Процесс перехода инфы из электрофизиологической памяти в структурную носит название консолидации (промежуточная память). В этот период происходит образование энграмм.
Выделяют произвольную, непроизвольную, образную и логическую.
Механизм формирования памяти: на первом этапе происходят электрофизиологические процессы, на втором биохимические реакции, а на третьем структурные изменения в стуктурах и синапсах ЦНС. Кратковременная память – элекрофизиологическая, промежуточная – нейрохимическая, долговременная – нейроструктурная. Следует выделить первичную и вторичную электрофизиологическую память.
Первичная электрофизиологическая память. Эта память кратковременна. Объем составляет 7+- единицы. Длительность хранения инфы составляет от нескольких секунд до 5 минут.
В основе механизма первичной электрофизиологической памяти лежит кратковременная циркуляция возбуждения благодаря кратковременной синаптической потенциации. Начальной стадией усвоения любой инфы является восприятие раздражителя. Инфа воспринимается рецепторами органов чувств и кодируется в виде рецепторного потенциала, который с помощью своего электрического поля обеспечивает возникновение нервных импульсов, предающихся в ЦНС.
Вторичная электрофизиологическая память.
Основой механизма этого вида памяти является долговременная циркуляция возбуждения благодаря долговременной синаптической потенциации. Одним из факторов, активирующим этот вид памяти являются эмоции. Длительность хранения информации определяется длительностью циркуляции возбуждения (10 – 15 часов).
Нейрохимическая (промежуточная) память. В данный период происходит консолидация памяти – перевод хранения инфы с помощью биохимических процессов в долговременную память. Консолидация продолжается не более четырех часов.
Нейроструктурная (долговременная) память. Ее основой является синтез белка и она не нарушается при экстремальных воздействиях на мозг. Формируется с помощью электрофизиологических и нейрохимических процессов. На данном этапе формирования памяти можно видеть структурные изменения в синапсах и отростках нейронов (ультраструктурная память), происходят незаметные молекулярные структурные изменения в нейронах (макромолекулярная память).
Образование условных рефлексов.
Условные рефлексы формируются не только на уровне неокортекса, но и на уровне мозжечка. Предполагают, что мозжечок контролирует точность выполнения движений во времени, т.к. только он обладает способностью в любой момент времени заблокировать любую двигательную реакцию или, наоборот, дать ей возможность реализоваться.
Основные правила выработки условных рефлексов. Необходимые условия:
Неоднократное совпадение во времени действия индифферентного раздражителя с безусловным.
Условный стимул должен предшествовать безусловному, на базе которого вырабатывается условный рефлекс.
Способствующие факторы:
Оптимальное состояние организма, в первую очередь состояние коры большого мозга
Активное функциональное состояние нервного центра безусловного рефлекса
Отсутствие посторонних сигналов из внешней среды и от внутренниихх органов
Оптимальное соотношение силы условного и безусловного раздражителей. Для образования и сохранения условных рефлексов существует определенный диапазон подкреплений: минимальный, оптимальный, максимальный. Чрезмерно сильный условный сигнал может препятствовать выработке условного рефлекса.
Основные стадии образования условного рефлекса.
Латентная стадия характеризуется выраженной концентрацией возбуждения в проекционных зонах коры условного и безусловного раздражителей и отсутствием условных поведенческих реакций.
Стадия генерализации условного рефлекса состоит в том, что условные реакции возникают на сигнальный и др раздражители (явление афферентной генерализации), а также в интервалах между предъявлениями условного стимула (межсигнальные реакции). В этот период различные биоэлектрические сдвиги широко распространены по коре и подкорковым структурам. В период генерализации наблюдается синхронизация биоэлектрической активности во многих участках коры и подкорковых образованиях – дистантная синхронизация биопотенциалов, в основе которой лежит процесс дивергенции возбуждения.
Стадия специализации обеспечивает дифференцировку стимулов, специализацию условно – рефлекторного навыка. Изменения биотоков более ограничены и приурочены к действию условного стимула.
Механизм образования условного рефлекса (временной связи).
На клеточном и молекулярном уровнях временная связь замыкается с помощью механизмов памяти.
В начале выработки условного рефлекса временная связь образуется только за счет механизмов электрофизиологической памяти – распространения возбуждения между двумя возбужденными корковыми центрами, возбуждаемыми в результате действия безусловного и условного раздражителей. Образованию функциональной связи между корковыми центрами ассоциируемых раздражителей способствует синхронизация биопотенциалов.
По мере действия условного и безусловного раздражителей и повторяющегося возбуждения соответствующих центров кратковременная память переходит в долговременную, происходят структурные изменения в нейронах.
Процессы включения механизмов памяти при замыкании временной связи осуществляются следующим образом. Временная связь образуется между корковым центром безусловного рефлекса и корковым центром сенсорной системы, на рецепторы которой действует условный раздражитель, т.е. связь замыкается в коре большого мозга. В основе замыкания временной связи лежат процессы дивергенции возбуждения в ЦНС и формирования доминантных очагов возбуждения. Импульсы, вызываемые индифферентным сигналов с любого участка кожи и др органов чувств, поступают в кору, где образуют очаг возбуждения и обеспечивают его дивергенцию. Если после индифферентного сигнала подается подкрепление, то в коре возникает более мощный второй очаг возбуждения и также наблюдается дивергенция возбуждения по коре. Неоднократное сочетание в опытах индифферентного и безусловного раздражителя облегчает прохождение импульсов от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса. Это явление называется синаптической потенциацией (проторение пути), оно ведет к формированию доминанты. Образование временной связи в коре Павлов назвал замыканием условно – рефлекторной дуги: подача только условного сигнала приводит к возбуждению коркового центра безусловного рефлекса, возникает условный рефлекс.
Образование условных рефлексов с прямыми и обратными связями является общей закономерностью ВНД. В процессе выработки условного рефлекса образуется двусторонняя связь между двумя центрами: корковым концом сенсорной системы, на рецепторы которого действует условный раздражитель, и центром безусловного рефлекса, на базе которого вырабатывается условный рефлекс.
Взаимосвязь левого и правого полушарий (перенос возбуждения из одного полушария в другое) при выработке условных рефлексов обеспечивается наличием комиссуральной системы (мозолистого тела, передней, задней, гиппокампальной и хабенулярной комиссур, межбугрового сращения), которая анатомически соединяет два полушария. Элементарные формы взаимодействия двух полушарий могут осуществляться через четверохолмие и РФ ствола.
В начале выработки условного рефлекса наблюдается дивергенция возбуждения в ЦНС, затем развивается синаптическая потенциация на пути между двумя центрами (проторение пути) и, наконец, формируются доминантные отношения между двумя центрами.
Учение И.П.Павлова о торможении условных рефлексов: виды торможения, условия их возникновения, примеры; механизм, биологическое значение.
Торможение условных рефлексов.
Различают врожденное и приобретенное торможение условных рефлексов.
Безусловное торможение подразделяется на два вида внешнее и запредельное.
Внешнее торможение заключается в ослаблении или прекращении наличного условного рефлекса при действии какого – либо постороннего раздражителя. Эту реакцию, возникшую на изменение внешней среды (рефлекс на новизну), Павлов назвал рефлексом «что такое?». Он состоит в настораживании и подготовке организма к действию на случай внезапно возникающей необходимости (нападение, бег и др.). с повторением действия дополнительного раздражителя реакция на этот сигнал ослабевает и исчезает, если организму не требуется предпринимать каких – то действий.
Механизм внешнего торможения: посторонний сигнал сопровождается появлением в коре большого мозга нового очага возбуждения, который при достаточной силе раздражителя оказывает угнетающее влияние на текущую условно – рефлекторную деятельность по механизму доминанты. Новый очаг возбуждения находится вне дуги наличного условного рефлекса.
Значение внешнего раздражения состоит в том, что оно способствует экстренному приспособлению организма к меняющимся условиям внешней и внутренней среды и дает возможность при необходимости переключиться на другую деятельность в соответствии с ситуацией.
Запредельное торможение возникает при действии чрезвычайно сильного условного сигнала.
Механизм запредельного торможения: между силой условного торможения и величиной ответной реакции имеется определенное соответствие – закон силы: чем сильнее условный сигнал, тем сильнее условно – рефлекторная реакция. Выполняет охранительную функцию.
Условное торможение. Локализовано в пределах нервного центра данного условного рефлекса. Различают четыре основных виды: угасательное, запаздывательное, дифференцировочное и условный тормоз.
Угасательное возникает при повторном применении условного сигнала и без подкрепления. Вначале условный рефлекс ослабевает, а затем полностью исчезает, но через некоторое время может восстановиться при возобновлении подкрепления. Скорость угасания зависит от интенсивности условного сигнала и биологической значимости подкрепления: чем они значительные, тем труднее совершается угасание.
Запаздывательное возникает при отставании подкрепления на 1 – 3 мин относительно начала действия условного сигнала. Постепенно возникновение условной реакции сдвигается к моменту подкрепления.
Дифференцировочное вырабатывается при дополнительном включении раздражителя, близкого к условному, и неподкреплении его.
Условный тормоз возникает при добавлении к условному сигналу любого другого раздражителя и неподкреплении этой комбинации.
Условное торможение можно ослабить и восстановить угасший ранее рефлекс, если во время проявления торможения подействовать посторонним сигналом – возникает ориентировочно – исследовательский рефлекс. Это явление называется растормаживанием.
Механизм условного торможения условных рефлексов изучен недостаточно. На клеточном уровне зарегистрирована гиперполяризации корковых нейронов. В процесс условного торможения широко вовлекаются различные мозговые структуры, при угасательном торможении амплитуда ВПСП постепенно снижается без заметного перехода в гиперполяризацию клеточной мембраны.
Торможение условных рефлексов можно объяснить тем, что при неподкреплении условного раздражителя доминантные взаимоотношения двух очагов возбуждения исключаются и остается только один очаг возбуждения, вызванный условным сигналом. Проторенный путь между доминантными очагами возбуждения постепенно устраняется, т.к. исчезает синаптическая потенциация, сформированная ранее.
Значение всех видов условного торможения заключается в устранении ненужной в данное время деятельности.
Иррадиация и концентрация возбуждения при выработке условных рефлексов, их индукционные взаимодействия (опыты И.П.Павлова на собаках с касалками). Аналитико-синтетическая деятельность коры больших полушарий, динамический стереотип, условно-рефлекторное переключение.
Основные правила выработки условных рефлексов. Необходимые условия: 1. неоднократное повторение во времени действия индифферентного раздражителя с безусловным; 2.условный стимул должен предшествовать безусловному.
Способствующие факторы: 1. Оптимальное состояние организма; 2. Активное функциональное состояние нервного центра безусловного рефлекса; 3. Отсутствие посторонних сигналов из внешней среды и от внутренних органов. 4. Оптимальное соотношение силы условного и безусловного раздражителей.
В процессе образования условного рефлекса выделяют три стадии.
Латентная стадия характеризуется выраженной концентрацией возбуждения в проекционных зонах коры условного и безусловного раздражителей и отсутствием условных поведенческих реакций.
Стадия генерализации условного рефлекса состоит в том, что условные реакции возникают на сигнальный и другие раздражители, а также в интервалах между предъявлениями условного стимула. В этот период различные биоэлектрические сдвиги широко распространены по коре большого мозга и подкорковым структурам. Наблюдается синхронизация биоэлектрической активности во многих участках коры и подкорковых образованиях – дистантная синхронизация биопотенциалов, в основе которой лежит процесс дивергенции возбуждения.
Стадия специализации обеспечивает дифференцировку стимулов, специализацию условно – рефлекторного навыка.
Множество раздражителей внешнего мира и внутренней среды организма воспринимаются рецепторами и становятся источниками импульсов, которые поступают в кору больших полушарий. Здесь они анализируются, различаются и синтезируются, соединяются, обобщаются. Способность коры разделять, вычленять и различать отдельные раздражения, дифференцировать их и есть проявление аналитической деятельности коры головного мозга.
Сначала раздражения анализируются в рецепторах, которые специализируются на световых, звуковых раздражителях и т. п. Высшие формы анализа осуществляются в коре больших полушарий. Аналитическая деятельность коры головного мозга неразрывно связана с ее синтетической деятельностью, выражающейся в объединении, обобщении возбуждения, которое возникает в различных ее участках под действием многочисленных раздражителей. В качестве примера синтетической деятельности коры больших полушарий можно привести образование временной связи, которое лежит в основе выработки условного рефлекса. Сложная синтетическая деятельность проявляется в образовании рефлексов второго, третьего и высших порядков. В основе обобщения лежит процесс иррадиации возбуждения.
Анализ и синтез связаны между собой, и в коре происходит сложная аналитико-синтетическая деятельность.
Динамический стереотип. Внешний мир действует на организм не единичными раздражителями, а обычно системой одновременных и последовательных раздражителей. Если система последовательных раздражителей часто повторяется, это ведет к образованию системности, или динамического стереотипа в деятельности коры головного мозга. Таким образом, динамический стереотип представляет собой последовательную цепь условно-рефлекторных актов, осуществляющихся в строго определенном, закрепленном во времени порядке и являющихся следствием сложной системной реакции организма на сложную систему положительных (подкрепляемых) и отрицательных (неподкрепляемых, или тормозных) условных раздражителей.
Условно-рефлекторное переключение. Понятие "условно-рефлекторное переключение" ввел Э.А.Асратян. Оно состоит в том, что один и тот же раздражитель может стать условным сигналом различных условных реакций. Физиологическим механизмом условно-рефлекторного переключения является образование комплексов разного сигнального значения из одних и тех же основных сигналов с добавлением разных обстановочных раздражителей.
Например, гудок в одной экспериментальной камере может быть сигналом пищевой реакции, а в другой камере - сигналом оборонительного рефлекса, или, один и тот же сигнал в первую половину дня может служить оборонительным условным раздражителем, а во вторую половину - пищевым сигналом. Из этого примера видно, что здесь имеет место образование разных условных рефлексов на комплексы раздражителей, общим компонентом которых служит сигнал, применяемый экспериментатором в качестве условного раздражителя (гудок). Мозг животного во время опыта анализирует составные части комплекса раздражителей, выделяет "основные" агенты и "переключатели". Он синтезирует, объединяет их и связывает с определенной деятельностью.
Учение И.П.Павлова о типах ВНД: критерии типологических различий, методы определения типов, характеристика типов, роль генотипа и среды в формировании фенотипа.
Критерии оценки свойств НС.
Сила процесса возбуждения отражает работоспособность нервной клетки. Она проявляется в функциональной выносливости, т.е. способности выдерживать длительное или кратковременное, но сильное возбуждение, не переходя в состояние охранительного торможения. Если слабое раздражение вызывает сильно возбуждение, ведущее к развитию охранительного торможения, то сила процесса возбуждения слабая, а тип НС высоковозбудимый.
Сила процесса торможения понимается как функциональная работоспособность НС при реализации условного торможения. Сила процесса торможения проявляется в способности к образованию различных тормозных условных реакций – угасания, дифференцировки или запаздывания.
Уравновешенность означает равенство силы процессов возбуждения и торможения, вовлеченных в выработку условных рефлексов или условного торможения.
Подвижность нервных процессов указывает на способность НС быстро реагировать на изменения среды и переходить от одних условных рефлексов к другим в зависимости от требований среды.
Павлов выделил 4 основных типа ВНД: 1. Сильный, уравновешенный, подвижный; 2. Сильный, неуравновешенный; 3. Сильный, уравновешенный, инертный; 4. Слабый.
Сильный, уравновешенный, подвижный = сангвиник. Такой человек решительный, энергичный, с высокой возбудимостью, подвижен. Сильный, уравновешенный, инертный = флегматик. Спокойный, медлительный, со слабы проявлением чувств. Сильный, неуравновешенный = холерик. Вспыльчивый, с высоким уровнем активности, энергичный. Слабый = меланхолик. Унылый, тоскливый, склонный к мрачным мыслям.
У человека тип ВНД определяют, во – первых, посредством выработки различных условных рефлексов и условного торможения; во – вторых с помощью специальных методик.
Существует два методических подхода определения силы нервных процессов: исследования изменения реакций при разной интенсивности раздражителя и при длительном раздражении. В последнем случае сила определяется кА устойчивость величины латентного периода сенсомоторной реакции на многократное повторение раздражителей средней величины. Один из вариантов заключается в следующем: испытуемому дается 100 световых раздражителей с интервалом 1 – 3 с, каждый раз при загорании лампочки испытуемый должен как можно сильнее нажать на рычаг. Показателем, характеризующим силы нервных процессов, будет среднее значение самой двигательной реакции на последние 20 раздражений.
Определение уравновешенности нервных процессов чаще всего проводится по количеству положительных двигательных реакций на тормозный раздражитель при выработке дифференцировки. Для этого испытуемому дается инструкция, не смотря на циферблат, останавливать стрелку на цифре 10. Высчитывают число точных, опережающих и опаздывающих реакций, вычисляют среднюю величину отклонений времени, вариационный размах и алгебраическую сумму отклонений.
Для исследования подвижности нервных процессов используют показатели длительности последействия условного тормоза на двигательные реакции, соотношение латентных периодов условных двигательных реакций до и после переделки стереотипа. Испытуемому дается задача при загорании зеленой лампочки нажимать рычажок вправо, при загорании красной влево. Подают 50 раздражителей. Затем дают обратную задачу. Также подают 50 раздражителей. Количественными характеристиками подвижности являются среднее время реакции и точность.
Роль генотипа в формировании ВНД.
Доказательства влияния наследственного фактора получены в различных опытах. Так, в опыте Рундквиста от популяции активных крыс через 12 поколений получились активные крысы, а от пассивных пассивные.
Влияние среды показано в опытах на щенках Майорова и Выржиковского. Одна группа животных содержалась в закрытых клетках, а вторая группа в свободных вольерах. У собак первой группы выработалось сильное пассивно – оборонительное поведение.
Эмоции: определение, классификация, состояние организма во время эмоций; причины возникновения, значение. Роль эмоциональных напряжений в возникновении неврозов.
Эмоции – ярко выраженные переживания организма в форме удовольствия или неудовольствия.
Классификация эмоций. В зависимости от степени выраженности выделяют: настроение – эмоция – аффект; по знаку: положительные и отрицательные. Отрицательные эмоции могут быть стеническими (повышение активности организма (физической, духовной) и астеническими (угнетение деятельности организма). Положительные обычно сопровождаются повышением деятельности. Выделяют следующие основные эмоции: радость, интерес, удивление, горе, отвращение, гнев, презрение, страх вина, стыд, наслаждение, сострадание.
Выделяют три вида эмоциональных процессов: аффекты, собственно эмоции и чувства.
Аффекты – сильные и относительно кратковременные эмоциональные переживания, сопровождающиеся резко выраженными двигательными и висцеральными проявлениями.
Собственно эмоции – длительно текущее состояние, иногда лишь слабо проявляющееся во внешнем поведении. Выражают личностное отношение к складывающейся или возможной ситуации.
Предметные чувства возникают как специфическое обобщение эмоций и связаны с представлением или идеей о некотором объекте, конкретном или отвлеченном. Выражают устойчивые эмоциональные отношения.
Настроение – течение или поток гедонически ориентированных идей, мыслей и образов, извлекаемых из памяти. Их объединяет общий положительный или отрицательный гедонический фон.
Состояние организма во время эмоций. Стеническая отрицательная эмоция обычно сопровождается возбуждением ЦНС, выбросом в кровь катехоламинов, ведущих к активации ряда систем организма – усилению деятельности ССС, дыхания, повышению тонуса мышц, двигательной активности. Деятельность ЖКТ при этом, как правило, угнетается. Изменения ЭЭГ , характерные для эмоций, наиболее отчетливо возникают в лобной области. У лиц с доминированием + эмоций регистрируется альфа ритм и медленные составляющие ЭЭГ, а у лиц с преобладанием гнева бета активность.
Причины возникновения эмоций. Выделяют три основные теории.
Биологическая теория (Анохин), в основу которой положена концепция функциональной системы: эмоция связана с появлением биологическое потребности, которая может сопровождаться – эмоцией, удовлетворения потребности, в результате чего возникает + эмоция.
Потребностно – информационная теория (Симонов), согласно которой в основе появления эмоций лежат потребности и информация, необходимая для ее достижения.
Если объем информации недостаточен для удовлетворения потребности, возникает – эмоция, если достаточен +, в результате удовлетворения потребности.
Представление Косицкого заключается в том, что для достижения цели необходимы определенная инфа, энергия и время: если существующие у организма инфа, энергия и время недостаточны, то возникает напряжение.
Значение эмоций.
Мобилизация физических и интеллектуальных ресурсов (эмоция) способствует сосредоточению внимания, обостряет мыслительную деятельность и чувствительность сенсорных систем, облегчает запоминание большого объема инфы и на более длительный срок. Вместе с тем отрицательная астеническая эмоция сопровождается демобилизацией ресурсов организма.
Коммуникативная роль осуществляется с помощью глосса, мимических пантомимических движений, позволяющим передавать свои переживания др людям.
+эмоции оказывают благотворное влияние на здоровье человека.
- эмоции обладают длительным последействием, способны суммироваться; могут при частых и повторных проявлениях переходить в застойную форму.
Кора головного мозга и внутренние органы: слово как лечебный и патогенный фактор (примеры), понятие о психосоматической медицине, механизм взаимодействия коры большого мозга и внутренних органов (кортиковисцеральные взаимоотношения).
Психосоматическая медицина – направление в медицине и психологии, занимающееся изучением влияние психогенных факторов на возникновение и течение соматических заболеваний, а заболеваний на психику.
Быков обнаружил интероцептивные сигналы, поступающие по афферентным проводникам в подкорковые ядра и некоторые отделы коры. На основе интероцептивной инфы, поступившей в кору большого мозга, могут образоваться временные связи, т.е. сигнализация от внутренних органов может включаться в общие механизмы условно – рефлекторной интегративной деятельности мозга. Эти обобщения послужили основой для создания теории кортиковисцеральных взаимоотношений. Нарушение этих взаимоотношений одна из причин психосоматических расстройств.
Нейрофизиологические модели психосоматических расстройств.
Ошибка нервных процессов возбуждения и торможения. Она происходит при одновременной активизации с помощью одного и того же условного раздражителя противоречащих друг другу рефлексов, например пищевого и оборонительного.
Образование патологических условных рефлексов.
Кортиковисцеральная теория Быкова и Курцина. Эта теория объяснила происхождение внутренних болезней следующим образом: кора большого мозга может непосредственно влиять на состояние внутренних органов (по механизму безусловных и условных рефлексов); следовательно, причиной психосоматических заболеваний может быть нарушение соотношения возбуждения и торможения на уровне коры большого мозга.
Экстероцептивный условный рефлекс можно образовать на деятельность любого внутреннего органа, функция которого не поддается произвольному управлению. Всякое колебание внешней среды по механизму временной кортикальной связи может изменить функцию висцеральных систем. Кора большого мозга может при определенных условиях воздействовать на работу висцеральных систем, на все их функции , вплоть до обменных.
Влияние коры большого мозга на внутренние органы осуществляется при участии лимбико – ретикулярной, вегетативной и эндокринной систем. Возникновение психосоматических заболеваний может быть связано с первичным нарушением корковых механизмов управления висцеральными органами, обусловленным перенапряжением возбуждения и торможения в коре большого мозга.
Пусковыми механизмами психосоматических расстройств могут быть факторы внешней и внутренней среды или нарушения экстеро и интероцептивной сигнализации, приводящие к конфликтной ситуации между возбуждением и торможением в коре и подкорке, что в конечном итоге вызывает невроз или психоз.
Кортиковисцеральный цикл – объединение инфы, поступающей в ЦНС от всех интеро и экстерорецепторов и от коры и др. структур мозга к внутренним органам. Это объединение происходит на уровне коры и др. нервных уровней, а также при участии эндокринных механизмов регуляции.
Особенности ВНД человека: назвать, дать определение, охарактеризовать в сравнении с ВНД животных. Осознаваемые и подсознательные реакции, и их значение.
ВНД – совокупность нейрофизиологических процессов, обеспечивающих сознание, подсознательное усвоение поступившей информации и обучение в онтогенезе всем видам деятельности, в том числе приспособительному поведению.
Целенаправленная планируемая трудовая деятельность человека включает предварительное планирование в виде схем, чертежей, расчетов, будущих испытаний продуктов труда. Человек активно изменяет окр мир, в соответствии с потребностями общества. Поведение животных направлено только на приспособление в природе.
Мышление человека принципиально отличается от мышления животных. Во – первых, мышление человека может осуществляться на основании инструкций. Во – вторых, у человека имеется образное (конкретно – предметное) и абстрактное (словесно – логическое) мышление, у животных только образное.
Павлов выделил три типа ВНД, присущих только человеку:
Художественный тип характеризуется преобладанием первой сигнальной системы.
Мыслительный тип характеризуется преобладанием второй сигнальной системы.
Средний тип.
Неосознаваемая (подсознательная) деятельность. На подсознательном уровне происходят следующие процессы.
Усвоение ранее поступившей инфы.
Восприятие подпороговых внешних раздражителей: на уровне подсознания можно выработать условные рефлексы.
Переработка импульсов от внутренних органов.
На уровне подсознания протекают хорошо отработанные, автоматизированные элементы работы, а также ходьба, бег и др привычная деятельность, глубоко усвоенные нормы поведения.
Деятельность мозга во сне направлена на переработку инфы, полученной до сна, на запоминание ее.
Интуиция связана с процессами творчества, которые не контролируются сознанием. И7туиция – источник гипотез, открытий, возможных благодаря трансформации и рекомбинации следов памяти.
Значение подсознательной деятельности мозга состоит в том, что ресурсы сознания во время подсознательной деятельности освобождаются и переключаются на другую деятельность; реакции, протекающие на уровне подсознания энергетически более выгодны, наиболее экономны.
Осознаваемая деятельность включает два вида.
Во – первых, к ней относятся все виды психической деятельности: ощущение, внимание, мышление, мотивация, эмоции.
Во – вторых, на уровне сознания происходит неавтоматизированная физическая работа. Она осуществляется в подавляющем большинстве случаев, т.к. при нарушении концентрации внимания немедленно совершаются ошибки в производственной деятельности.
Значение правого и левого полушарий в интегративной деятельности мозга. Сигнальные системы: понятие о первой и второй сигнальной системах человека, формирование их в онтогенезе, развитие второй сигнальной системы в антропогенезе, взаимоотношения первой и второй сигнальной систем.
Изолированное левое полушарие также эффективно обеспечивает владение письменной и устной речью, как и нерасщепленный мозг.
Изолированное правое полушарие способно к зрительному или тактильному распознаванию форм и в определенной степени к пониманию речи.
Леве полушарие воспринимает и обрабатывает преимущественно сигналы второй сигнальной систем (абстрактное мышление), а правое преимущественно сигналы первой сигнальной системы (образное мышление). Левое полушарие обрабатывает инфу последовательно, аналитически, расчленено, по принципу индукции (от частного к общему), лучше воспринимает абсолютные признаки предметов и временные отношения. Правое полушарие обрабатывает инфу одновременно, синтетически, по принципу дедукции (от общего к частному), при этом лучше воспринимаются пространственные и относительные признаки предметов.
Эмоции. Левое полушарие обусловливает в основном положительные эмоции, контролирует проявление более слабых эмоций. Правое полушарие обусловливает преимущественно отрицательные эмоции, контролирует проявление сильных эмоций.
Речевые центры. У большинства людей центры речи располагаются в левом полушарии, у 1 – 5 % в правом. Левое полушарие больше отвечает за смысл речи, правое за интонацию.
Речевые функции у правшей локализованы в левом полушарии. У 70 % леворуких центр речи расположен в левом полушарии, у 15 % в правом, у 15 % и в левом, и в правом.
Сигнальные системы – совокупность систем организма, обеспечивающих формирование сигналов; сами сигналы для передачи инфы др особям; совокупность структур, воспринимающих подобную инфу от др особей. Сигналами ПСС являются элементы языка животных, свойственного и человеку. Сигналами ВСС являются специфические элементы языка человека.
Речь в антропогенезе сформировалась как средство общения в процессе охоты на диких зверей, защиты при их нападении, сооружении жилища и т.д. Вначале это были отдельные звуки в виде сигналов. Затем отдельные звуки превратились в более сложные сигналы, образовались слоги, слова, фразы.
ПСС – совокупность структур организма, воспроизводимых и воспринимаемых ими сигналов и самих сигналов, обеспечивающих обмен инфы между особями с помощью элементов языка животных. ВСС - совокупность структур организма, воспроизводимых и воспринимаемых ими сигналов и самих сигналов, обеспечивающих обмен инфы между людьми с помощью специфичных элементов языка человека.
Сон: определение, фазы, механизм, значение, виды, оценка глубины сна с помощью ЭЭГ (неглубокий, средней глубины и глубокий сон). Сновидения и факторы, побуждающие их возникновение. Физиологические основы гипнотических состояний, значение их для клиники.
Сон – циркадный биоритм пониженной активности мозга, обеспечивающий отдых организма, при котором выключены сознание и механизмы поддержания естественной позы, снижена чувствительность сенсорных систем.
Засыпанию способствует ряд факторов: соблюдение режима сна (циркадный биоритм), утомление нервных клеток, ослабление активности сенсорных систем, удобная поза.
Признаки сонливости: снижение уровня сознания; зевание; понижение чувствительности сенсорных систем; урежение сердцебиения и дыхания, снижение секреторной деятельности желез.
Виды сна: 1. Естественный: периодический суточный и периодический сезонный; 2. Искусственный: наркотический и гипнотический; патологический: при анемии мозга, поражении структур мозга.
Различают две фазы сна: медленную и быструю. Обычно вначале развивается фаза медленного сна (ортодоксальный сон, 80 % ночного сна), затем быстрого сна (парадоксальный сон, 20 %), фазы чередуются.
Медленный сон по мере углубления проходит 4 стадии, прерываясь стадией парадоксального сна. В ЭЭГ в это время выделяется медленная электрическая активность в виде синхронизации (частота волн уменьшается, амплитуда возрастает). На первой стадии преобладают альфа волны, на второй тета волны, на третьей появляются сонные веретена и К- комплексы (серия медленных потенциалов большой амплитуды) в сочетании с высоковольтными дельта волнами, на четвертой стадии преобладают дельта волны.
Быстрый сон наступает после четвертой стадии медленного сна. На ЭЭГ появляется электрическая активность, характерная для бодрствующего состояния – десинхронизация (учащение волн и снижение их амплитуды); наблюдаются характерные монофазные спайки с частотой 60 – 70 в мин; возникают сновидения.
Сновидения – возникающие во сне образы объектов и явлений, которые в действительности могут не существовать.
Сновидения встречаются в обоих фазах сна, но чаще сопровождают парадоксальный сон. Происхождение сновидений объясняют активацией следов долговременной памяти.
Среди факторов, побуждающих сновидения, выделяют раздражители, действующие на организм во время сна; предшествующая сну деятельность; избыточная импульсация от переполненных или больных внутренних органов; биологические потребности.
Значение сна.
Сон обеспечивает отдых организма.
Играет роль в процессах метаболизма. Медленный сон помогает восстановлению внутренних органов, поскольку через гипоталамус либерины воздействуют на гипофиз, способствуя выделению гормонов роста, который участвует в биосинтезе белков в периферических тканях. Парадоксальный сон восстанавливает пластические свойства нейронов головного мозга, усиливает процессы в клетках нейроглии, которые обеспечивают нейроны питательными веществами и кислородом.
Сон – приспособление организма к изменению освещенности.
Сон способствует усвоению инфы.
Механизмы бодрствования и сна.
В настоящее время известно две теории сна: гуморальная и нервная.
Гуморальная теория. Из мочи и спинномозговой жидкости выделен фактор S, вызывающий медленноволновый сон. Существует и фактор сна с БДГ. Выделен пептид дельта сна, индуцирующий глубокий сон. ВИП и бета – эндорфин углубляют состояние сна. Тиреотропный гормон прерывает сон.
Нервная теория сна. Электрическое раздражение преоптической области гипоталамуса вызывает синхронизация электроэнцефалограммы и поведенческий сон. Обратный эффект наблюдается при раздражении заднего гипоталамуса.
Разрушение ядер шва устраняет синхронизация ЭЭГ и медленный сон. Нейроны ядер шва синтезируют серотонин и направляют его через свои аксоны к РФ, гипоталамусу, лимбической системы. Устранение синтеза серотонина у кошки устраняет медленный сон, сохраняя лишь парадоксальный сон.
Стимуляция голубого пятна (средний мозг, медиатор – норадреналин) вызывает торможение нейронной активности во многих структурах мозга при росте двигательного возбуждения животного и ЭЭГ – десинхронизации. Полагают, что активирующее действие голубого пятна осуществляется посредством торможения тормозных интернейронов. Разрушение ядер шва ведет к полной бессоннице в течение нескольких дней, но за несколько последующих недель сон нормализуется. Двустороннее разрушение голубого пятна ведет к полному исчезновению БДГ фаз, не влияя на медленноволновой сон.
В каудальной части мозгового ствола обнаружены структуры, отвечающие за медленный сон. Наличие сходных гипногенных структур было обнаружено и в задней части моста. Двигательные и ЭЭГ проявления фазы парадоксального сна связаны с активацией структур в области моста.
Важную роль в цикле сон – бодрствование играет РФ ствола мозга. Высокочастотное раздражение этой структуры ведет к мгновенному пробуждению. Рядом с голубым пятном имеется группа гигантских ретикулярных нейронов активность которых важна в о время парадоксального сна.
В составе холинергической системы мозга различают активирующие и тормозные структуры. Уровень активности холинергических нейронов увеличивается во время бодрствования и парадоксального сна.
Гипноз – искусственно вызванное с помощью внушения состояние человека, при котором не полностью выключено сознание и снижены реакции на все раздражители, кроме слов гипнотизирующего, к которым человек становится крайне восприимчив. В ответ на монотонные или многократные стимулы непременно развивается внутреннее торможение. Если такая стимуляция продолжается, то наступает сон.
Переходный период между бодрствованием и сном назван гипнотическим состоянием. Выделяют три фазы гипнотического состояния: уравнительную, парадоксальную, ультрапарадоксальную. В парадоксальной фазе сильные раздражители вызывают слабую реакцию, а слабые сильную. В ультрапарадоксальной реакция может быть получена посредством неадекватного стимула.
Основным физиологическим условием внушаемости является снижение тонуса коры больших полушарий и возникающая при этом функциональная расчлененность корковой деятельности. Парадоксальная фаза – сильные раздражители внешнего мира уступают слабым стимулам, идущим от слов гипнотизера.
В стадии гипноза волны ЭЭГ сходны с ЭЭГ бодрствования.
Стадии гипноза.
Гипноидность – сопровождается психическим и мышечным расслаблением, миганием и закрыванием глаз
Легкий транс – развивается кателепсия конечностей (конечности могут долгое время находиться в необычном положении)
Средний транс – возникает неполная амнезия, изменения личности, возможны простые гипнотические внушения.
Глубокий транс – полная амнезия, сомнамбулизм, фантастические внушения.
При гипнозе изменяется состояние коры больших полушарий, которое меняет соотношение первой и второй сигнальными системами. Снижается контроль второй сигнальной системы над первой и облегчается воспроизведение пережитых состояний и зрительных образов. Возбуждение коры при восприятии слов гипнотизера оказывается изолировано от влияния остального мозга; воздействия на него становятся чрезвычайно сильными продолжительными последействиями, что объясняет длительный эффект гипнотерапии.
Известны два вида гипнотерапии – посредством гипноза и под гипнозом. В терапии посредством гипноза наиболее распространены два подхода: снятие симптомов путем словесного внушения и перестройка личности (замещение симптомов). В терапии под гипнозом выделяют: метод прямого внушения с целью изменения поведения человека; метод катарсиса, позволяющий проявить подавленные эмоции, что дает возможность выявить причину психосоматических расстройств; метод гипноанализа, заключающийся в активации симптомов под гипнозом и их последующем анализе в состоянии бодрствования или под гипнозом.
Целенаправленное поведение: социально и биологически детерминированное поведение (мотивации), особенности труда в условиях современного производства, утомление и активный отдых (И.М.Сеченов), биоритмы и утомление. Классификация биоритмов, их устойчивость и изменчивость, десинхронозы.
Мотивации – побуждение организма к действию с целью с целью удовлетворения потребности.
Мотивации подразделяют на три группы: биологические, социальные и идеальные. Их также делят на первичные и вторичные.
Сложные целенаправленные формы поведения возникают на основе специфических внутренних состояний, обусловленных возникновением той или иной потребности. Таки внутренние состояния Павлов назвал влечениями организма, сейчас их называют мотивациями.
Мотивационное возбуждение, возникнув в центрах гипоталамуса вследствие непосредственного влияния на них факторов внутренней среды или же в результате поступления к ним импульсаций от различных органов и тканей, постепенно начинает распространятся на структуры лимбической системы нейронные комплексы передних долей коры большого мозга. Дальнейшее усиление активности в гипоталамусе через таламо – тегментальные связи стимулирует восходящую РФ среднего мозга, что приводит к генерализованному возбуждению всех отделов коры большого мозга, предопределяя новый доминантный статус мотивационного возбуждения. С этого момента все механизмы мозга по принципу взаимодействия объединяются в единый комплекс, направляющий организм к удовлетворению той потребности, на базе которой сформировалась доминантная мотивация.
Гипоталамические центры являются ведущими в развитии мотивационного поведения, поэтому их называют пейсмекерными центрами. Разрушение этих центров устраняет возможность возникновения мотивации.
В области вентромедиальных ядер гипоталамуса и в латеральных его отделах обнаружены нейроны, чувствительные к содержанию в крови глюкозы. В области супраоптических и паравентрикулярных ядер гипоталамусы обнаружены нейроны, чувствительные к уровню осмотического давления крови и т.д. электрическая стимуляция этих ядер вызывает соответственно питьевые, пищевые мотивации, мотивации страха и агрессии.
Поиск, завершение целенаправленного поведения организма во внешней среде, приводящие к удовлетворению потребности, осуществляются корой большого мозга, которая оказывает непрерывные нисходящие влияния на инициативные центры гипоталамуса. Удовлетворение потребности ведет к прекращению мотивационного возбуждения.
Все мотивации обладают выраженной нейрофизиологической и нейрохимической специфичностью. Так, мотивация страха блокируется альфа – адреноблокаторами, пищевые мотивации холинолитиками.
Мотивационные возбуждения вызывают актуализацию механизмов памяти, тем самым повышая вероятность достижения полезного результата на основе ранее приобретенного опыта. Мотивации активируют сенсорные системы. Любая мотивация повышает активность моторной системы.
Утомление – временное снижение умственной или физической работоспособности, вызванное предшествующей деятельностью и сопровождаемое желанием прекратить работу или снизить величину нагрузки. При этом уменьшается мышечная сила и выносливость, появляется масса лишних движений, возрастает количество ошибочных действий, изменяется ЧСС и частота дыхания, увеличивается АД, ухудшается мышление, увеличивается время зрительно – моторных реакций. Ослабляется внимание – его устойчивость и переключаемость, выдержка, настойчивость, снижаются возможности памяти.
Стадии утомления: первая – ощущение усталость выражено незначительно, производительность труда практически не снижена; вторая – ощущение усталости выражено ярко, производительность труда снижена существенно; третья – производительность труда может быть снижена до нулевых показателей, ощущение усталости выражено сильно, сохраняется после отдыха, иногда становится хроническим.
Степени переутомления начинающееся, легкое, выраженное и тяжелое. Оно характеризуется снижением творческой активности, умственной и физической работоспособности, нарушением биоритмов и ростом заболеваемости.
Сеченов обнаружил явление активного отдыха: деятельность одних мышечных групп или конечности способствует устранению утомления, возникающего при работе других мышечных групп или конечности. Активный отдых – отдых, заполненный другим видом деятельности.
Биоритмы обычно классифицирует по частоте осцилляции (длительности периодов). По этому признаку выделяют биоритмы с высокой частотой (период 0,5 ч), ритмы средней частоты (0,5 – 28 ч), циркасептанные (около 7 суток), макроритмы (20 суток – 1 год), мегаритмы (более 10 лет). Мегаритмы не могут считаться биоритмами: в организме они сами по себе не возникают, а представляют собой реакции на процессы, периодически возникающие на солнце.
Удобнее классифицировать биоритмы по их происхождению: физиологические, геофизические и геосоциальные.
Физиологические биоритмы – непрерывная циклическая деятельность всех органов, систем, отдельных клеток организма обеспечивает выполнение их функций.
Физиологические биоритмы генетически запрограммированы, обладают видовой специфичностью.
Значение физиолгических ритмов заключается в обеспечении нормального функционирования клеток, органов и систем организма. Возможность быстрого изменения частоты физиологических ритмов обесепечивает приспособительные реакции – адаптацию организма к различным условиям жижизнедеятельности. Нередко это мгновенные приспособительные реакции. Например, после начала физической нагрузки очень быстро (через 1 – 2 сек) повышаются частота и сила сердечных сокращений, интенсивность дыхания, что улучшает доставку кислорода и других питательных веществ к усиленно работающим мышцам. Деятельность ЖКТ при этом угнетается. В покое физиологические ритмы быстро возвращаются к исходному уровню.
Геофизические биоритмы – циклические изменения деятельности клеток, органов систем и организма в целом, а также резистентности, миграции и размножения, обусловленные геофизическими факторами. Геофизические биоритмы представляют собой циклические колебания физиологических биоритмов, обусловленные изменениями факторов среды обитания. Главными геофизическими биоритмами являются околомесячные и окологодовые. Генетически запрограммированы, не обладают видовой специфичностью.
Геофизические биоритмы сформировались под влиянием природных факторов, во многом связаны с временами года, фазами луны. Таковыми являются сезонные изменения цвета шерсти, циклические изменения обмена веществ.
Значениие геофизических биоритмов заключается в обеспечении приспособления организма к циклическим изменениям в природе.
Геосоциальные биоритмы. Формируются под влиянием социальных и геофизических факторов. Главными геосоциальными биоритмами являются околосуточные (циркадианные) и околонедельные (циркасептанные), длительностью около 7 – 8 дней. Установление биоритма является результатом реализации наследственной инфы. Геосоциальные биоритмы, когда они уже сформированы, сохраняются в условиях изоляции организма.
Циркадианные биоритмы отличаются стабильностью и строгой закономерностью. Они синхронизированы с вращением земли вокруг своей оси, длительность их периода 20 – 28 часов.
Значение геосоциальных биоритмов заключается в приспособлении организма к режиму труда и отдыха.
Десинхроноз – расстройства в организме, возникающие вследствие рассогласования между сложившимися циркадианными биоритмами и новым режимом труда и отдыха.
Защитные системы организма и их проявления в различных условиях деятельности человека. Понятие о перекрестной резистентности и сенсибилизации организма.
Адаптивные реакции реализуются на клеточном, органном, системном и организменном уровнях. Все адаптивные механизмы объединяют в две группы по двум основным критериям: в зависимости от времени их становления в онтогенезе – врожденные и приобретенные; по скорости возникновения и длительности действия: срочные, долговременные и постоянные. Срочные включаются сразу после начала действия безусловных или условных раздражителей.
Врожденные и приобретенные адаптивные механизмы.
Наследственны (видовой иммунитет) – невосприимчивость одного вида к микроорганизмам, вызывающим заболевания у других видов.
Прямая (специфическая) резистентность – повышение устойчивости организма к длительно или кратковременно, но повторно действующему раздражителю.
Перекрестная (неспецифическая) резистентность – повышение устойчивости не только к действующему, но и к другому или нескольким раздражителям.
Стрессор – любое сильное физическое или психогенное воздействие на организм, оказывающее часто отрицательное влияние - дистресс, реже положительное – эустресс. При действии стрессора возникает состояние напряжения организма – стресс. Стрессовая реакция включает следующую триаду: увеличение коркового слоя надпочечников и повышение его активности, уменьшение вилочковой железы и лимфатических желез, точечные кровоизлияние и кровоточащие язвочки в слизистой оболочке желудка и кишечника. Однако патологических изменений в организме может и не наблюдаться, если стрессор недостаточно силен, а устойчивость высока.
При длительном действии стрессора возникает общий адаптационный синдром. Он включает в себя три стадии: тревоги, резистентности и истощения.
Реакция тревоги обычно возникает в первые 6 ч от начала действия стрессора и длится 1 – 2 суток. Поток афферентных импульсов, возникших вследствие действия стрессора, поступает в ЦНС. В рУсловно-рефлекторное переключение. Понятие "условно-рефлекторное переключение" ввел Э.А.Асратян. Оно состоит в том, что один и тот же раздражитель может стать условным сигналом различных условных реакций. Физиологическим механизмом условно-рефлекторного переключения является образование комплексов разного сигнального значения из одних и тех же основных сигналов с добавлением разных обстановочных раздражителей.
Например, гудок в одной экспериментальной камере может быть сигналом пищевой реакции, а в другой камере - сигналом оборонительного рефлекса, или, один и тот же сигнал в первую половину дня может служить оборонительным условным раздражителем, а во вторую половину - пищевым сигналом. Из этого примера видно, что здесь имеет место образование разных условных рефлексов на комплексы раздражителей, общим компонентом которых служит сигнал, применяемый экспериментатором в качестве условного раздражителя (гудок). Мозг животного во время опыта анализирует составные части комплекса раздражителей, выделяет "основные" агенты и "переключатели". Он синтезирует, объединяет их и связывает с определенной деятельностью. В реакцию вовлекается гипоталамо – гипофизарно – надпочечниковая система: повышается выброс катехоламинов и кортикостероидов, в ЦНС повышается метаболизм РНК, белков и др веществ. Возрастает расход энергии. Обычно возникают ярко выраженные переживания (эмоции), они чаще отрицательные, в обмене веществ преобладает диссимиляция. Вначале срабатывают срочные адаптивные механизмы.
В фазу резистентности все перечисленные симптомы исчезают благодаря формирования адаптивных механизмов.
Стадия истощения развивается при длительном и чрезмерном действии стрессора, когда истощаются активные механизмы.
В основе резистентности лежат клеточные механизмы адаптации и формирование соответствующей функциональной системы. Каждый раз при действии стрессора остается структурный след. Структурные следы – основа долговременного повышения резистентности организма. Элементами механизма увеличения резистентности при длительном действии стрессора являются увеличение синтеза РНК и белков, мобилизация иммунной системы, увеличение выработки глюкокортикоидов, которые стимулируют глюконеогенез, увеличение работы гамма – глобулинов – все это обеспечивает прямую и перекрестную резистентности.
Деадаптация – постепенно уменьшение и полное исчезновение адаптации в случае прекращения действия фактора. Процесс деадаптации обычно развивается в течение 1- 2 месяцев.
Адаптация у физической нагрузке. В ее основе лежит активация синтеза мышечных белков. Увеличивается количество иРНК, что приводит к повышению числа рибосом и полисом, в которых происходит синтез белка. Систематическая двигательная деятельность приводит к увеличению емкости капиллярной сети в мышцах, содержания гликогена, АТФ, креатинфосфата, дыхательных ферментов.
Возрастающее кол – во метахондрий способствует возрастанию способности мышц утилизировать пируват. При этом ограничивается накопление молочной кислоты обеспечивается возможность мобилизации жирных кислот, повышается способность к интенсивной и длительной мышечной работе. Увеличивается толщина моторных нервных волокон, кол – во терминальных нервных веточек, усиливается синтез белка не только в мышцах, но и в других органах.
Статический режим деятельности мышц приводит к более глубоким перестройкам сосудистой системы и нервных окончаний: капилляры изменяют ход, идут не параллельно мышечным волокнам, а оплетают их; аксоны нейронов двигательных единиц делятся на большее число терминалей, подходящих к мышечным волокнам. Надежность функционирования ОДА возрастает за счет увеличения поперечника трубчатых костей и утолщения их компактного вещества.
Рационально дозируемые нагрузки способствуют повышению возможностей ССС и дыхательной системы, системы крови и ЦНС. Увеличение работоспособности сердца связано с активацией работы кальциевого насоса в эндоплазматической сети, увеличением кол – ва митохондрий в кардиомиоцитах, активацией ферментов, ответственных за транспорт субстратов окисления.
Систематический физические упражнения делают организм боле устойчивым
к заболеваниям, повышают устойчивость организма к недостатку О2,
улучшают утилизацию О2 с помощью активации ферментативных систем.
Физические тренировки обеспечивают экономное расходование энергии,
улучшают координационную деятельность ЦНС. Вследствие движений
улучшается функциональное состояние ЦНС за счет проприоцептивной
импульсации от мышц. Активируются и уравновешиваются возбудительные
и тормозные процессы. В клетках коры большого мозга увеличивается
содержание РНК, имеющей непосредственное отношение к механизмам
памяти. Усиливаются ассоциативные процессы и творческая деятельность.
В головном мозге увеличивается кол – во капилляров и их длина. Двигательная активность снижает скорость склерозирования сосудов, поскольку снижается уровень холестерина в крови.
Адаптация и деадаптация к гипокинезии. Вначале включаются реакции, активирующие недостаток двигательной активности: возбуждается симпатоадреналовая система в связи с эмоциональным напряжением, в крови увеличивается кол – во кортикостероидов. Гипокинезия сильно уменьшает нагрузку на сердце. Однако на первых порах работа сердца усиливается за счет возбуждения симпатической НС, выброса в кровь катехоламинов, сужения сосудов и повышения АД. Вследствие активации симпатоадреналовой системы возрастает катаболизма в тканях, что ведет к большему употребления О2 и вентиляции легких. Однако эти реакции угасают при продолжительной гипокинезии, следствием которой становится снижение катаболических процессов: понижается потребление организмом О2, в крови уменьшается содержание углекислоты, молочной кислоты, ослабевают дыхание и деятельность сердца, снижается кровяное давление.
Уменьшается масса скелетной и сердечной мышц, снижается их энергетический потенциал.
Абсолютная вынужденная обездвиженность может закончиться гибелью, т.к. сопротивляемость организма очень низкая. Вместо фазы резистентности идет медленное истощение всех функций.
При относительной гипокинезии устанавливается низкоэнергетический гемостазис – фаза резистентности. Но она отличается нестабильностью, резким снижением общей резистентности организма, предрасположенностью к любым патологическим процессам вследствие недостаточности функционирования различных систем организма.
Своевременная активация мышечной деятельности постепенно нормализует функции всех систем организма.

