

7 семестр / Ингибиторы-Общий путь катаболизма-1
.pdf220
Лекция. Игибиторы дыхательной цепи. Коэффициент фосфорилирования. Окислительное декарбоксилирование пирувата
Ингибиторы и разобщители окисления и фосфорилирования
Ингибиторы ферментов цепи переноса электронов и окислительного фосфорилирования
Ферментные комплексы |
|
Ингибиторы |
|
|
|
||
Комплекс I (НАДН-дегидрогеназа) |
Ротенон (инсектицид) |
||
|
|
Барбитураты |
(амобарбитал, |
|
|
секобарбитал) |
|
|
|
Пиерицидин А (антибиотик) |
|
|
|
|
|
Комплекс |
III |
антимицин А (антибиотик) |
|
(убихинолдегидрогеназа) |
|
|
|
|
|
|
|
Комплекс IV (цитохромоксидаза) |
CO, CN-, H2S, N3- |
|
|
Комплекс V (АТФ-синтаза) |
|
олигомицин (антибиотик) |
|
|
|
дициклогексилкарбодиимид |
|
|
|
|
|
АТФ/АДФ-транслоказа |
|
атрактилозид |
|
|
|
|
|
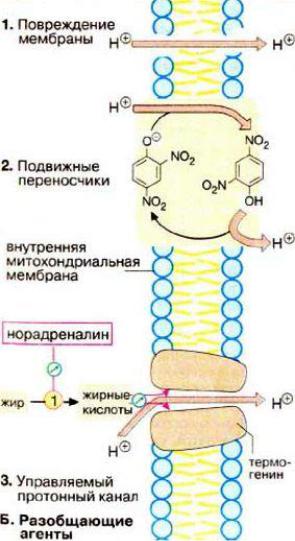
Разобщение окисления и фосфорилирования
Некоторые вещества разобщают окисление и фосфорилирование. Например, 2,4-динитрофенол:
OH |
|
- |
|
O |
|
NO |
+ |
NO |
2 |
- H |
|
|
2 |
|
|
+ |
|
|
+ H |
|
NO |
|
NO |
2 |
|
|
|
|
2 |
Это гидрофобное вещество легко диффундирует через митохондриальную мембрану как в ионизированной, так и в неионизированной форме и, следовательно, может переносить ионы водорода через мембрану в сторону их меньшей концентрации. Поэтому 2,4- динитрофенол уничтожает электрохимический потенциал митохондриальной мембраны, а энергия рассеивается в форме теплоты. Потребление кислорода и окисление субстратов при этом продолжается, но синтез АТФ невозможен.
221
Поскольку энергия окисления при разобщении рассеивается, то разобщители повышают температуру тела (пирогенное действие).
Коэффициент фосфорилирования (Р/О)
При окислении субстратов, идущих с участием НАД-зависимых дегидрогеназ происходит включение трех молей фосфата в АДФ и образования 3 моль АТФ в расчете на 1/2 моль потребляемого кислорода О2
(Р/O = 3)/
При окислении субстратов через флавиновые дегидрогеназы образуется 2 моль АТФ (Р/О = 2).
ЭНЕРГЕТИЧЕСКИЙ ОБМЕН И ТЕПЛОПРОДУКЦИЯ
Превращение энергии пищевых веществ в организме включает следующие этапы:
аккумуляция в НАДН или ФАДН2 высокоэнергетических электронов; превращение в форму электрохимического потенциала митохондриальной мембраны; аккумуляция в АТФ;
использование АТФ для совершения работы.

222
На всех этапах трансформации часть энергии рассеивается в форме теплоты:
теплота |
|
теплота |
теплота |
НАДН |
+ |
АТФ |
Работа |
Δμ H |
В состоянии покоя расходование энергии на внешнюю работу минимально и теплопродукция становится главным путем расхода энергии. В частности, значительный вклад в образование теплоты вносят транспортные АТФ-азы. Например, Na+-K+-АТФаза. Такое состояние энергетического обмена называют основным обменом. Интенсивность теплового обмена можно оценить по величине теплопродукции. Для взрослого человека она равна примерно 350 кДж/ч (2000 ккал за сутки). Калорийность потребляемой пищи должна быть равна этим тратам.
Характеристика общего пути катаболизма
К общему пути катаболизма относятся окислительное декарбоксилирование пировиноградной кислоты и цикл лимонной кислоты (ЦЛК). Некоторые специфические пути вливаются в общий путь на стадии пирувата, другие - на стадии ацетил-КоА. Ряд веществ вступает в общий путь катаболизма на промежуточных стадиях ЦЛК. Именно в общем, пути катаболизма образуется основная масса первичных доноров водорода для дыхательной цепи, хотя они образуются и в специфических путях катаболизма.
Окислительное декарбоксилирование пировиноградной кислоты
. Общая характеристика пируватдегидрогеназного комплекса
При окислении глюкозы, глицерина и аминокислоты аланина образуется пировиноградная кислота (пируват). Пируват далее может быть окислен до СО2 в общем пути катаболизма. Реакции промежуточного обмена протекают в цитоплазме, а окисление пирувата - в митохондриях, поэтому пируват транспортируется в митохондрии специальным переносчиком через мембрану. Первый шаг катаболизма пирувата - окислительное декарбоксилирование.
Вэтом процессе участвуют ферменты, работающие в определенной последовательности и объединенные в мультиферментный
пируватдегидрогеназный комплекс (ПДГК).
Всостав этого комплекса входит 3 фермента и 5 коферментов. Ферменты:
Пируватдегидрогеназа (Е1) Дигидролипоилацетилтрансфераза (Е2)
Дигидролипоилдегидрогеназа (Е3).

223
Коферменты:
Тиаминдифосфат (ТДФ) - производное витамина В1
|
NH |
|
|
H |
|
|
|
|
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
N |
CH |
N |
S |
|
O |
O |
|
2 |
|
|
||||
|
|
|
|
CH |
CH |
O P O P OH |
|
H C |
N |
H C |
2 |
2 |
|
|
|
3 |
|
3 |
|
|
OH |
OH |
|
|
|
|
|
|
|
||
Никотинамидадениндинуклеотид (НАД+) - производное витамина никотинамида РР
Флавинадениндинуклеотид (ФАД) - производное витамина В2 Кофермент А (КоА-SH)
Липоевая кислота ковалентно связана с боковой цепью лизина фермента Е2 (липоамид)
|
|
O |
|
CH |
C |
|
2 |
|
S |
S |
NH |
1. |
CH |
|
- |
|
+ |
+ E |
|
ТДФ |
||
|
C COO |
+ H |
1 |
|||||||
|
|
3 |
|
|
|
|
|
|
||
|
|
|
|
O |
|
|
|
|
|
|
|
|
пируват |
|
|
|
|
|
|
||
2. |
E |
1 |
ТДФ CH CH |
+ |
|
|
E2 |
|||
|
|
|
|
3 |
|
|
|
|
||
|
|
|
|
OH |
|
|
|
S |
|
S |
|
|
|
|
|
|
|
окисленный |
|||
|
|
|
|
|
|
|
липоамид |
|||
3. |
|
|
|
E |
+ |
KoA SH |
||||
|
|
|
|
2 |
||||||
|
|
|
S HS |
|
|
|
|
|
|
|
|
O |
C |
|
|
|
|
|
|
|
|
|
|
|
CH |
|
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
4. |
|
|
|
E2 |
+ E3 |
ФАД |
|
|||
|
|
|
|
|
||||||
|
|
SH |
SH |
|
|
|
|
|
|
|
5. |
|
E |
3 |
ФАДН |
+ |
НАД |
+ |
|
|
|
|
|
|
|
|||||||
|
|
|
2 |
|
|
|
|
|
|
|
(CH ) |
E |
2 |
|
|
|
|
|
|
||
|
2 4 |
|
|
|
|
|
|
|
||
Lys |
|
|
|
|
|
|
|
|
|
|
CO |
|
|
|
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
E |
1 |
ТДФ CH CH |
|
|||||
|
|
|
|
|
|
|
3 |
|
||
|
|
|
|
|
|
|
OH |
|
||
|
|
гидроксиэтилтиамин- |
|
|||||||
|
|
дифосфат |
|
|
||||||
|
|
E |
1 |
ТДФ + |
E |
2 |
||||
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
S HS |
|
|
|
|
|
|
|
|
|
O |
C |
|
|
|
|
|
|
|
|
|
|
CH |
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
ацетиллипоамид |
|||
|
|
|
|
|
E |
2 |
+ KoA S С CH |
|||
|
|
|
|
|
|
|
|
|
3 |
|
SH |
|
|
SH |
|
|
|
O |
|
||
восстановленный |
|
|||||||||
ацетил-КоА |
||||||||||
липоамид |
|
|
|
|
|
|
||||
|
|
|
|
E |
2 |
+ E |
3 |
ФАДН |
|
|
|
|
|
|
|
|
|
2 |
|
||
S |
S |
|
|
|
|
|
|
|
|
|
E3 |
ФАД + |
|
|
|
+ |
|
||||
НАДН + Н |
|
|||||||||
Две последние реакции необходимы для регенерации активных форм коферментов, участвующих в окислительном декарбоксилировании пирувата. Образовавшийся в пятой реакции НАДН поступает в дыхательную

224
цепь под действием НАДН-дегидрогеназы. При окислении в дыхательной цепи 1 моль НАДН образуется 3 моль АТФ.
Суммарное уравнение окислительного декарбоксилирования пирувата:
CH |
- |
+ KoA SH + НАД |
+ |
KoA S С CH |
+ НАДН + CO |
C COO |
|
||||
3 |
|
|
|
3 |
2 |
|
O |
|
|
O |
|
пируват |
|
|
ацетил-КоА |
|
|
Регуляция ферментов пируватдегидрогеназного комплекса
Ингибирование продуктами реакции:
Ацетил-КоА и НАДН подавляют активность ПДГ-комплекса. Регуляция нуклеотидами по принципу обратной связи.
Пируватдегидрогеназа ингибируется ГТФ и активируется АМФ. Регуляция путем ковалентной модификации.
Пируватдегидрогеназа дезактивируется при фосфорилировании по остатку серина киназой пируватдегидрогеназы, при этом образуется неактивная фосфорилированная форма фермента (ПДГ-b) . При дефосфорилировании, катализируемом фосфотазой, активность фермента восстанавливается (ПДГ- a). Ацетил-КоА и НАДН являются положительными эффекторами киназы пируватдегидрогеназы. НАДН и ацетил-КоА накапливаются при увеличении клеточного заряда, поэтому увеличение уровня АТФ также приводит к увеличению активности киназы. При увеличении концентрации пирувата, АДФ, КоА-SH, НАД+ активность киназы снижается, а, следовательно, возрастает количество активной формы ПДГ-а.
10.6. Значение окислительного декарбоксилирования пирувата
Реакции пируватдегидрогеназного комплекса связывают метаболические пути гликолиза, глюконеогенеза, синтеза жирных кислот с циклом лимонной кислоты. ПДГ-комплекс имеет огромное значение в поддержании гомеостаза. Энергетический метаболизм аэробных тканей (например, мозга) очень зависит от превращения пирувата в ацетил-КоА, поэтому такие ткани более чувствительны к нарушениям ПДГ-комплекса. Большинство нарушений, связанных с недостаточностью пируватдегидрогеназной активности, обусловлены мутациями пируватдегидрогеназы. Главными последствиями таких нарушений являются энцефалопатии и церебральные молочные ацидозы.