Новіков А., Барабаш-Красни Б. Сучасна систематика рослин. Загальні питання
.pdf539

540
231. Нектарники (закінчення)
Інколи |
серед |
інтрафлоральних |
або |
симплікатній |
структурних зонах |
||||||
нектарників розрізняють nectaria caduca |
гінецея; |
|
|
|
|
||||||
(нектарники, що сполучені з опадаючими |
б) зону об’єднаного септального |
||||||||||
елементами квітки або ж гомологічні до |
нектарника – представлена сполученими |
||||||||||
них) та nectaria persistentia (нектарники, |
в центрі зав’язі камерами у єдину спільну |
||||||||||
що |
сполучені |
з |
неопадаючими |
порожнину, яка може розміщуватися в |
|||||||
елементами квітки або ж гомологічні до |
гемісинасцидіатній або гемісимплікатній |
||||||||||
них) (Smets 1986; Smets et al. 2000). |
структурних зонах гінецея; |
|
|||||||||
Окремо |
слід |
відзначити |
септальні |
в) зону вивідних каналів нектарника |
|||||||
нектарники |
– |
нектарники |
деяких |
– представлена ізольованими каналами, |
|||||||
однодольних рослин, що представлені |
щовиконуютьфункціювиведеннянектару |
||||||||||
екзогенними порожнинами (камерами), |
на поверхню зав’язі і, залежно від форми |
||||||||||
розташованими в перегородках (септах) |
нектарника, можуть розміщуватися в |
||||||||||
зав’язі зрослоплодолисткової маточки. |
різних структурних зонах гінецея. |
||||||||||
При цьому нектар може виділяти лише |
|
|
|
|
|
|
|||||
внутрішня поверхня |
камер септальних |
Існують й інші підходи до класифікації |
|||||||||
нектарників, де він накопичується та |
інтрафлоральнихісептальнихнектарників |
||||||||||
вивільняється назовні через канали. Або |
(Daumann 1970; Schmid 1985; Böhme |
||||||||||
жсекреторнуфункціюможутьвиконувати |
1988). |
|
|
|
|
|
|||||
також ділянки, які розташовані назовні. |
Smets & Cresens (1988) пропонують |
||||||||||
Simpson (1993) виокремлює три типи |
вживати |
замість |
вузького |
терміну |
|||||||
септальних нектарників за положенням |
септальні |
нектарники більш |
широкий |
||||||||
відносно гнізд зав’язі: а) інфралокулярні |
термін |
гіноплевральні |
нектарники, |
||||||||
– розташовуються нижче від гнізд зав’язі; |
що охоплює всі типи септальних |
||||||||||
б) інтерлокулярні – розташовуються на |
нектарників, зокрема і так звані зовнішні, |
||||||||||
рівні гнізд зав’язі; в) супралокулярні – |
які |
розташовуються |
на |
зовнішніх |
|||||||
розташовуються вище від рівня гнізд |
стінках зав’язі в деяких Magnoliidae, що |
||||||||||
зав’язі. |
|
|
|
|
додатково доводить |
спорідненість цієї |
|||||
|
|
|
|
|
|
групи рослин з однодольними. |
|
||||
У |
структурі септальних нектарників |
Насамкінець, |
нектар-продукуючу |
||||||||
Одінцова (2013) виділяє три основні |
функцію можуть переймати на себе й |
||||||||||
структурні зони: |
|
|
|
інші структури: а) гідатоди (наприклад, |
|||||||
а) зону роздільного септального |
у Fritillaria L.), що зазвичай виконують |
||||||||||
нектарника–представленаізольованими |
функцію виведення надлишку води з |
||||||||||
одна від одної камерами в перегородках |
рослини; б) елайофори (наприклад, у |
||||||||||
зав’язі, що можуть розміщуватися нижче |
деяких Malpighiaceae Juss.), які зазвичай |
||||||||||
від рівня гнізд зав’язі, у синасцидіатній |
продукують олії. |
|
|
|
|||||||
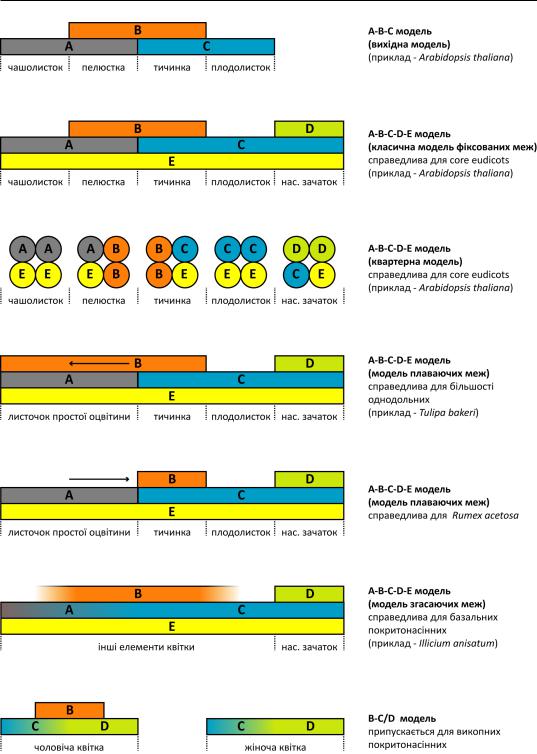
Рис. 197. Класифікація інтрафлоральних нектарників за розмірами, положенням у зав’язі, розташуванням вивідного каналу, а також структурою та розташуванням секреторної тканини (пунктир) за Daumann (1970). a-k – при гіпогінії; l-n – при семі-епігінії; o-v – при епігінії.
541

542
232. Квіткові формули
Існує багато різних способів для опису |
A2+2/Sto2+A2 означає, що у квітці може |
|||||||||||||
різноманіття квіток. Зокрема, це можуть |
міститися чотири тичинки по дві у двох |
|||||||||||||
бути текстові описи й ілюстрації, але |
колах, або ж дві з них можуть бути |
|||||||||||||
окрім них широко застосовують квіткові |
представлені стерильними стамінодіями. |
|||||||||||||
формули та діаграми. |
|
|
|
|
Для |
прикладу |
можна |
навести |
||||||
Квіткова формула (формула квітки) – |
порівняно |
|
просту |
формулу |
||||||||||
цечисельно-літерналінійнаформула,яка |
® K5 [C(5) A5] G(2), що описує |
|||||||||||||
описує симетрію квітки, а також кількість, |
радіальносиметричну квітку представників |
|||||||||||||
послідовність і |
характер |
розташування |
родини Boraginaceae з 5 вільними |
|||||||||||
елементів у квітці. |
|
|
|
|
|
чашолистками, 5 зрослими пелюстками, до |
||||||||
Квіткові |
|
формули |
|
|
можуть |
яких приросло 5 вільних тичинок, а також |
||||||||
бути |
традиційними |
текстовими |
верхньою зав’язю з 2 зрослих плодолистків |
|||||||||||
(використовують |
|
виключно |
|
текстові |
(Leins&Erbar2010). |
|
|
|
||||||
й |
математичні |
символи) |
або |
Більш складний приклад – формула |
||||||||||
напівграфічними (окрім текстових і |
® K(2):1↓ C1:2↓ [A30+10:2↓ Ĝ(3)*]Vp∞, |
|||||||||||||
математичних символів, використовують |
що описує резупіновану квітку орхідеї |
|||||||||||||
різні графічні елементи, в основному |
Cypripedium calceolus L.здвомазрослими |
|||||||||||||
для |
позначення |
характеру |
зростання |
й одним вільним чашолисткоми, що |
||||||||||
елементів квітки). Одні з найкращих |
формуютьзигоморфнучашечку;1вільною |
|||||||||||||
напівграфічних квіткових формул подано |
губою і 2 вільними бічними пелюстками, |
|||||||||||||
в книзі «Plant systematics: A phylogenetic |
що формують |
також зигоморфний |
||||||||||||
approach» (Judd et al. 1999). |
|
|
віночок; трьома повністю редукованими |
|||||||||||
Порядок розташування позначень у |
тичинками одного кола, а також однією |
|||||||||||||
квітковій формулі: а) стать квітки (якщо |
повністю редукованою тичинкою і двома |
|||||||||||||
потрібно); б) симетрія квітки; в) кількість, |
видозміненими тичинками другого кола, |
|||||||||||||
співвідношення |
і |
зростання |
елементів |
які формують зигоморфний андроцей; |
||||||||||
оцвітини; г) кількість, |
співвідношення |
нижньою |
|
радіальносиметричною |
||||||||||
та зростання тичинок; д) тип зав’язі, |
зав’яззю з трьох зрослих плодолистків, |
|||||||||||||
кількість, співвідношення і зростання |
що містять численні насінні зачатки з |
|||||||||||||
плодолистків; е) тип плода. |
|
|
паріетальною |
плацентацією |
(Prenner |
|||||||||
Кожен з елементів квітки у квітковій |
et al. 2010). Зверніть увагу, що символи, |
|||||||||||||
формуліпозначаютьзадопомогоюунікальної |
якіописуютьсиметрію,стоятьпіслячисла |
|||||||||||||
абревіатури або символу. Після кожної |
елементів окремих груп, а не перед |
|||||||||||||
абревіатури |
зазначають |
число |
відповідних |
їхніми абревіатурами. Цю саму формулу |
||||||||||
елементівіхарактерїхзростання. |
|
можна записати у спрощеному вигляді: |
||||||||||||
Також |
позначають |
|
усі |
|
можливі |
↓ K(2):1 C1:2 [A2 Ĝ(3)]. |
|
|
|
|||||
або принаймні основні варіанти у |
Детальніше |
з |
принципами |
і |
||||||||||
варіабельностічислаелементівквіткитаїх |
проблемами ужитку |
квіткових |
формул |
|||||||||||
редукції. Наприклад, фрагмент формули |
можна ознайомитися в статтях Prenner |
|||||||||||||
A6-∞ означає, що тичинок може бути |
et al. (2010) та Ronse De Craene et al. |
|||||||||||||
від 6 до багатьох. А фрагмент формули |
(2014). |
|
|
|
|
|
||||||||


545

546
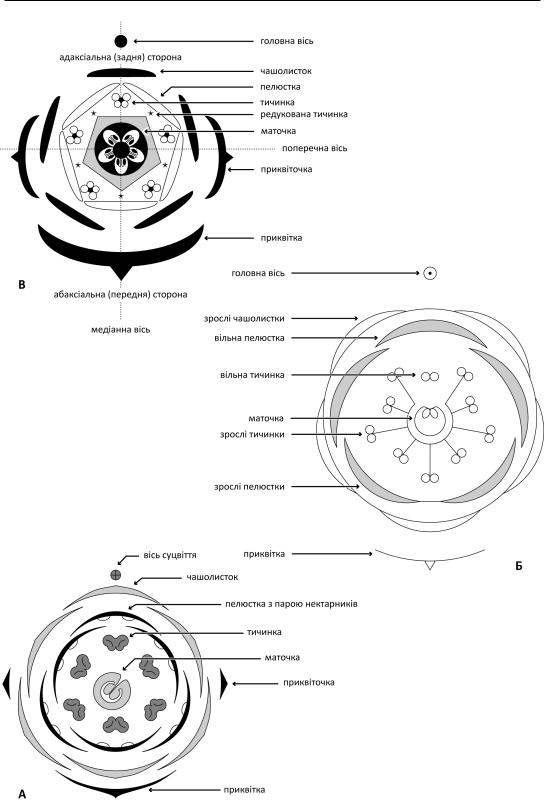
234. A-B-C-D-E модель ідентичності елементів квітки
Молекулярно-генетичні |
дослідження |
п’яти основних типів. Саме MADS- |
||||||
є потужним підґрунтям для розвитку |
комплекси |
регулюють експресію генів, |
||||||
сучасної ботаніки, а їхні результати |
які відповідають за розвиток тих чи інших |
|||||||
широко |
застосовують |
для |
вирішення |
елементівквітки. |
|
|||
найрізноманітніших |
проблем, |
які |
A-B-C-D-E |
|
модель |
чудово |
||
неможливо було розв’язати раніше. |
виправдовувала себе для рослин із |
|||||||
Зокрема,1991рокунапідставідослідження |
квітками, що мають подвійну оцвітину, а |
|||||||
Arabidopsis thaliana (L.) Heynh. було |
експресія гомеотичних генів має чіткі й |
|||||||
запропоновано A-B-C модель ідентичності |
визначенімежі(модельфіксованихмеж). |
|||||||
елементівквітки(Coen&Meyerowitz1991), |
Проте закономірно постало питання, як |
|||||||
згідно з якою формування чашолистків, |
A-B-C-D-E модель реалізується в рослин із |
|||||||
пелюсток, |
тичинок |
і |
плодолистків |
простоюоцвітиною.Буловстановлено,що |
||||
контролюєтьсятакзванимигомеотичними |
в більшості однодольних експресія генів |
|||||||
генами (названі так, тому що можуть |
класу B відбувається в обох колах простої |
|||||||
спричиняти гомеозис – заміщення одних |
оцвітини, а також у тичинках. Водночас |
|||||||
елементів іншими, наприклад, коли на |
виявилося, що в Rumex acetosa L., який |
|||||||
місці пелюсток розвиваються тичинки). |
також має просту оцвітину, експресія генів |
|||||||
Було запропоновано розділяти гомеотичні |
класуBвідбуваєтьсявиключновтичинках. |
|||||||
гени на три класи – A, B і C. Експресія лише |
Це дало поштовх для розвитку моделі |
|||||||
генівкласуAіндукуєрозвитокчашолистків. |
плаваючих (зміщених) меж експресії |
|||||||
Експресія генів класів A і B – розвиток |
гомеотичних генів. Щобільше, виявилося, |
|||||||
пелюсток. Експресія генів класів B і C – |
що в базальних покритонасінних, які |
|||||||
розвиток тичинок. Експресія лише генів |
відрізняються |
значним різноманіттям |
||||||
класуC–розвитокплодолистків. |
|
організації квітки, експресія гомеотичних |
||||||
Проте |
подальші |
дослідження на |
генів й інших класів (A і С) проявляється |
|||||
інших рослинах засвідчили, що участь у |
по-різному |
та |
в різному |
діапазоні |
||||
програмі |
формування |
елементів |
квітки |
(модель згасаючих меж). Ця гіпотеза |
||||
беруть й інші два класи гомеотичних генів |
пояснює, зокрема, наявність перехідних |
|||||||
–DтаE.ЕкспресіягенівкласуDобумовлює |
морфологічних рядів між тичинками й |
|||||||
формування насінних зачатків, тоді як гени |
листочками оцвітини (Theissen et al. 2002; |
|||||||
класу E виступають як допоміжні. A-B-C |
Zahnet al. 2005a, 2005b). |
|
||||||
модель була модифікована в A-B C D-E |
Цікаво, що у древніх покритонасінних, |
|||||||
модель ідентичності |
елементів |
квітки |
найімовірніше, гомеотичні гени класу B |
|||||
(Theissen 2001; Theissen & Saedler 2001). |
відповідали за визначення статі, тобто за |
|||||||
Згодом було встановлено, що гомеотичні |
розвиток чоловічих (експресія генів класу |
|||||||
гени кодують фактори |
транскрипції |
B відбувається) або жіночих (експресія |
||||||
(специфічні протеїни, що, у свою чергу, |
не відбувається) квіток. Водночас генів |
|||||||
впливають на транскрипцію ДНК) класу |
класу A у цих рослин не було, що дає |
|||||||
MADS. Було запропоновано квартерну |
підстави відносити їх до моделі B-C/D |
|||||||
модель, згідно з якою MADS-протеїни |
(Erbar 2007, 2010). |
|
||||||
формують |
тетрамерні |
MADS-комплекси |
|
|
|
|
||