

62. Пути нейтрализации аммиака. Орнитиновый цикл.
В клетках происходитит связывание аммиака с образазованием нетоксичных соединений, кот-ые выводятся из организма с мочой (р-ции обезвреживания аммиака.). В разных тканях и органах обнаружено несколько типов таких р-ций.
1.Р-циея
связывания аммиака, протекающей во всех
тканях: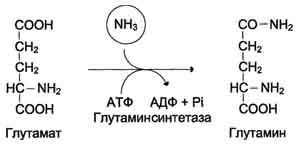 Глутамин
легко транспортируется через клеточные
мембраны путём облегчённой диффузии
и поступает из тканей в кровь, затем в
кишечник и почки. Основными
тканями-поставщикам глутамина служат
мышцы, мозг и печень.
Глутамин
легко транспортируется через клеточные
мембраны путём облегчённой диффузии
и поступает из тканей в кровь, затем в
кишечник и почки. Основными
тканями-поставщикам глутамина служат
мышцы, мозг и печень.
2. В клетках кишечника под действием фермента глутаминазы происходит гидролитическое освобождение амидного азота в виде аммиака
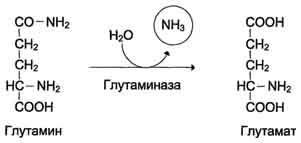
3.В почках также проис-ит гидролиз глутамина под дейс-ем глутаминазы с образ-ем аммиака. Глутаминаза почек значительно индуцируется при ацидозе, образ-щийся аммиак нейтрализует кислые продукты обмена и в виде аммонийных солей экскретируется .Эта р-ция защищает организм от излишней потери ионов Na+ и К+, которые также могут использоватся для выведения анионов и утрачиваются.
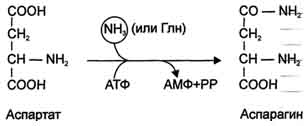
4. синтез аспарагина под действием аспарагинсинтетазы
5. Наиболее знач-ые кол-ва аммиака обезвреживается в печени путём синтеза мочевины.
Орнитиновый цикл(ОЦ)
ОЦ в печени выполняет 2 функции:
-превращение азота аминокислот в мочевину, которая экскретируется и предотвращает накопление токсичных продуктов( аммиака);
-синтез аргинина
Мочевина – осн-ой конечный продукт азотистого обмена, в составе кот-го из орг-зма выд-ся до 90% всего выводимого азота. Экскреция мочевины в норме сост-ет ∼25 г/сут.
Реакции синтеза мочевин:
Связывание аммиака в печени с диоксидом углерода с образ-ем карбамоилфосфата под дейст-ем карбамоилфосфатсинтетазы I
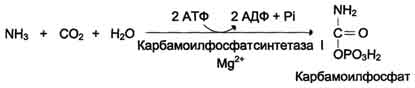
2. Под действием орнитинкарбамоилтрансферазы карбамоильная группа карбамоилфосфата переносится на α-аминокислоту орнитин, и образуется другая α-аминокислота – цитруллин
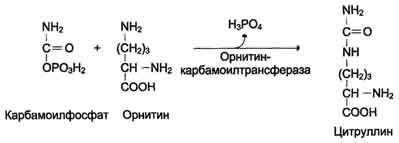
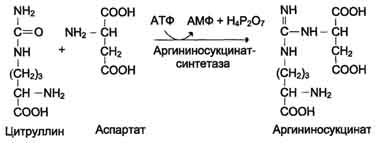
3. связывание цитруллин с аспартатом и образ-ие аргининосукцинат (аргининоянтарную к-ту)
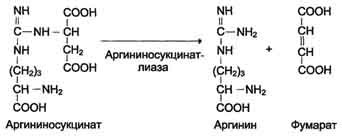
4. Далее фермент аргининосукцинатлиаза (аргининосукциназа) расщепляет аргининосукцинат на аргинин и фумарат
5. Аргинин подвергается гидролизу под действием аргиназы, при этом образуется орнитин и мочевина. Кофакторами аргиназы является ионы Са2+ или Мn2
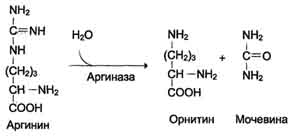
6. орнитин взаимодействует с новой молекулой карбамоилфосфата, и цикл замкнулся.
Суммарное уравнение синтеза мочевины:
СО2 + NH3 + Аспартат + 3 АТФ + 2 Н2О → Мочевина + Фумарат + 2 (АДФ + Н3Р04) + АМФ + H4P2O7.
63. Трансляция. Локализация процесса, стадии, необходимые условия. Активация аминокислот.
Трансляция - перевод генетического текста иРНК в последовательность АК в белке. Этапы трансляции:
1. Активация аминок-т;
2. инициация «начало»;
3. эллонгация «удлинение»;
4. терминация «прекращение»;
5. процессинг «созревание» и форм третичной структуры.
Место синтеза белка-рибосомы: прокариот:днк- 10^4, РНК-65%; эукариот – ДНК-10^5, РНК-50%. Прокариоты 70S→(Mg) 1) 50S:5S РНК, 28S РНК, 34S белка; 2) 30S: 15 РНК и 21 белок. Для каждой своя аминок-та: для серина, лейцина, аргенина-по 6Т-рнк, для аланина, треонина-по 4 Т-РНК. Т-рнк выполняет ф-цию адаптеров. Они акцепторным концом взаимодействуют с аминокислотами, а антикодоном - с кодоном мРНК. Аминоацил-тРНК синтетазы- катализирует р-цию специф связывания одной из 20 аминок-т с соответствующей тРНК. Ионы магния- Кофактор, стабилизир структуру рибосом. Белковые факторы инициации, элонгации, терминации-необхдля процесса трансляции (12 ф-ров инициации: elF; 2 ф-ра элонгации: eEFl, eEF2, и ф-ры терминации: eRF). Три последовательные химические р-ции приводят к включению аминок-ты в полипептидную цепь строящегося белка: 1 р-ция - активация аминок-ты за счет образ ангидридной связи между ее карбоксилом и фосфатом адениловой к-ты - и 2 р-ция - акцептирование аминоацильного остатка молекулой тРНК с образ сложноэфирной связи между карбоксилом аминок-ты и гидроксилом концевого рибозного остатка тРНК - происх внерибосомы, и обе р-ции катализируется ферментом аминоацил-тРНК-синтетазой. Синтез начинается с метионина у эукариот и формилметионина- у прокариот.