48. Окисление непредельных жирных кислот и жирных кислот с нечетным числом атомов углерода.
Окисл. ненасыщ. ЖК. Двойные связи природных ненасыщ. ЖК (олеиновой, линолевой и т.д.) имеют цис-конфигурацию, а в КоА-эфирах ненасыщ. кислот, являющихся промежуточными продуктами при β-окислении насыщ. ЖК, двойные связи имеют трансконфигурацию. Кроме того, последовательное удаление двууглеродных фрагментов при окисл. ненасыщ. В тканях есть фермент, кот. осущ. перемещение двойной связи из положения 3-4 в положение 2-3, а также изменяет конфигурацию двойной связи из цис- в транс-положение. При β-окислении ЖК, им-их две и более ненасыщ. связи, нужен еще один дополнител. фермент - 3-гидроксиацил-КоА-эпимераза.
В липидах многих растений и некот. морских организмов присутствуют ЖК с нечетным числом ат. углерода. Также у жвачных животных при переваривании углеводов в рубце образуется большое кол-во пропионовой кислоты, кот. содержит три углеродных атома. Пропионат всасывается в кровь и окисл. в печени и др. тканях. Установлено, что ЖК с нечетным числом углеродных атомов окисляются таким же образом, как и ЖК с четным числом углеродных ат., разница в том, что на последнем этапе расщепления (β-окисления) образуется одна молекула пропионил-КоА и одна молекула ацетил-КоА, а не 2 молекулы ацетил-КоА.
49. Синтез жирных кислот. Синтетаза жирных кислот.
Высшие ЖК могут быть синтезированы в организме из метаболитов углеводного обмена. Исходным соед-ем для этого биосинтеза явл. ацетил-КоА, образующийся в митохондриях из пирувата – продукта гликолитического распада глюкозы. Место синтеза жирных кислот – цитоплазма клеток, где имеется мультиферментный комплекс синтетаза высших жирных кислот. Этот комплекс сост. из 6 ферментов, связ-ых с ацилпереносящим белком. конечным продуктом явл. пальмитиновая кислота. Начал. р-цией синтеза ЖК явл. карбоксилирование ацетил-КоА с образованием малонил-КоА. Фермент ацетил-КоА-карбоксилаза активи-ся цитратом и ингиб-ся КоА-производными ВЖК.
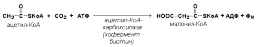
Затем ацетил-КоА и малонил-КоА взаимод. с SH-группами ацилпереносящего белка (рис.2).

Далее происходит их конденсация, декарбоксилирование и восстановление образовавшегося продукта (рис.3).

Продукт реакции взаимодействует с новой молекулой малонил-КоА и цикл многократно повторяется вплоть до образования остатка пальмитиновой кислоты.
Основные особенности биосинтеза ЖК по сравнению с β-окислением:
синтез жирных кислот в осущ. в цитоплазме клетки, а окисл. – в митохондриях;
участие в процессе связывания СО2 с ацетил-КоА;
в синтезе ЖК принимает участие ацилпереносящий белок, а в окисл. – коэнзим А;
для биосинтеза ЖК необходимы окислительно-восст. коферменты НАДФН, а для β-окисления – НАД+ и ФАД.
50. Биосинтез триглицеридов и фосфолипидов.
Биосинтез триглицеридов и фосфолипидов происходит в цитоплазме клеток. Первые реакции этих метаболических путей совпадают и включают образование фосфатидной кислоты.
Предшественники фосфатидной к-ты - глицерол и ЖК - вступают в р-цию в активной форме.
Образование активной формы глицерола (глицерол-3-фосфата) может происходить двумя путями (рис.1).

Рис.1. Пути образования глицерол-3-фосфата.
Первая реакция ацилирования глицерол-3-фосфата осуществляется за счёт КоА-производного насыщ. ЖК, напр., пальмитиновой. Образуется сложноэфирная связь в 1-м полож.
Вторая реакция ацилирования идёт, как правило, за счёт КоА-тиоэфира ненасыщ. ЖК, напр., олеиновой
В процессе синтеза триглицеридов происходит дефосфорилирование фосфатидной кислоты при помощи фосфатидатфосфатазы
В дальнейшем 1,2-диацилглицерол ацилируется третьей молекулой ацил-КоА, кот. может содержать остаток как ненасыщ., так и насыщ. ЖК
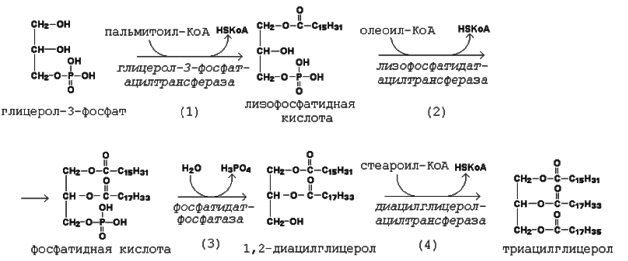
Рис.2. Реакции биосинтеза триацилглицеролов.
Фосфолипиды явл. производными фосфатидной кислоты и одного из аминоспиртов (напр., этаноламина, холина или серина). Р-ции образования фосфатидной кислоты (см. рис.2).
Аминоспирты, участвующие в синтезе фосфолипидов, вступают в р-цию в активной форме – в виде соед. с цитидиндифосфатом (ЦДФ).

Рис.3. Реакции образования активной формы этаноламина.
Далее остаток фосфоэтаноламина с ЦДФ-этаноламина переносится на 1,2-диацилглицерол, образующийся в результате дефосфорилирования фосфатидной кислоты (рис.4, р-ции 1 и 2).

Рис.4. Реакции синтеза фосфатидилхолина.
Синтез фосфатидилхолина происходит путём трёхкратного метилирования фосфатидидэтаноламина.