Сигнальные системы
Сигналинг (сигнальная трансдукция) – это передача сигнала рецептор → трансдуктор → эффектор. Сигнальные системы – это обязательный компонент организации многоклеточных организмов, так как взаимодействие клеток между собой происходит с помощью сигнальных молекул и их специфических сигнальных путей, которые координируют все биохимические процессов. Передача сигнала внутри клеток осуществляется с помощью слаженного каскада реакций, которые обеспечивают специфические ферментативные системы. Для некоторых механизмов сигнальных систем имеются внутриклеточные посредники первичного сигнала – вторичные мессенджеры.
Первичные сигнальные молекулы (лиганды):
Гормоны
Цитокины
Нейромедиаторы
Факторы роста
Рецепторы
Рецепторы (R)
локализация рецептора зависит от природы лиганда:
1. Гомоны белково-пептидной природы и гидрофильные сигнальные молекулы имеют специфический рецептор на мембране, а они в свою очередь бывают: метаботропные - семисегментные трансмембранные рецепторы 7 ТМS (сопряжённые с G-белками) и односегментарные трансмембраные рцепторы 1 ТМС, а также ионнотропные (ионные каналы).
Семисегментные трансмембранные рецепторы 7 ТМS :
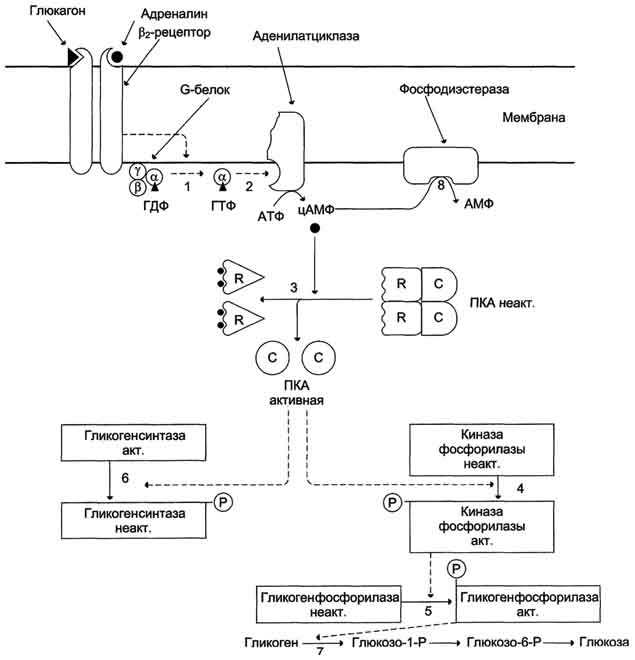
Rs рецепторы (стимулирующие) – активируют Gs белок и фермент аденилатциклазу: β-адренорецепторы/гормон адреналин, V2 -рецептор/гормон вазопрессин.
Ri рецепторы (ингибирующие) – активируют Gi белок, уменьшают активность аденилатциклазы: α2- адренорецепторы/гормон адреналин, М2-холинорецепторы/нейромедиатор ацетилхолин.
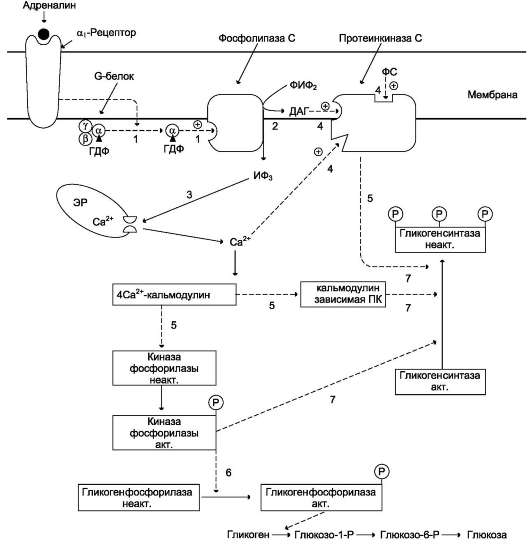
Rq Gq - фосфолипазы С: М1 и М2- холинорецепторы/нейромедиатор ацетилхолин, α1- адренорецепторы/гормон адреналин.
G-белки — это семейство белков, относящихся к ГТФ-азам, в своём сигнальном механизме используют замену ГДФ на ГТФ как молекулярный функциональный «выключатель».
гетеротримерные, большие (состоят из трёх субъединиц: α, β и γ)
малые (из одной полипептидной цепи: Ras, Rho, Arf и др.) У всех гетеротримерных G-белков сходный механизм активации:
активируются при взаимодействии со специфическими рецепторами, сопряженными с G-белками, при этом обменивая ГДФ на ГТФ
распадаясь на α и βγ-субъединицы
α-субъединица, связанная с ГТФ, воздействует на эффекторный фермент
инактивация происходит в результате медленного гидролиза ГТФ до ГДФ, после чего происходит реассоциация субъединиц α и βγ
Односегментные трансмембранные рецепторы ( 1 ТМS):
рецепторы, обладающие тирозинкиназной активностью, на них действуют: инсулин, инсулиноподобный фактор роста, цитокины
рецепторы, обладающие иной, не тирозинкиназной активностью, например, серин- и гуанилатциклазной и т.п., на них действуют: атриопептид, NO
Лиганд-зависимые ионные каналы:
Н-холинорецепторы для нейромедиатора ацетилхолина являются Na/K-каналами.
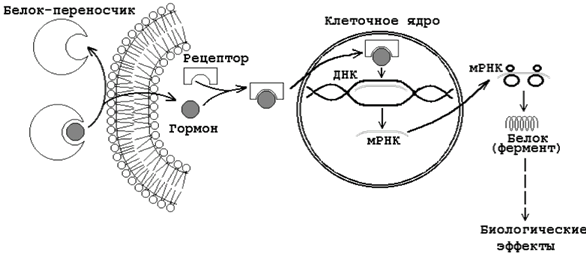
2. Небольшие липофильные сигнальные молекулы способны проникать в клетку за счёт диффузии. Рецепторы таких гормонов обычно являются растворимымицитоплазматическими (цитозольными) или ядерными белками.
Цитозольные – рецепторы к стероидным гормонам (женские и мужские половые гормоны, кальцитриол, гормоны коры надпочечников)
Ядерные – на них действуют гормоны щитовидной железы (T3, тироксин).
Ядерные рецепторы гормонов представлены семейством негистоновых белков, связанных с хроматином ядра клетки.
Вторичные мессенджеры
Вторичные мессенджеры
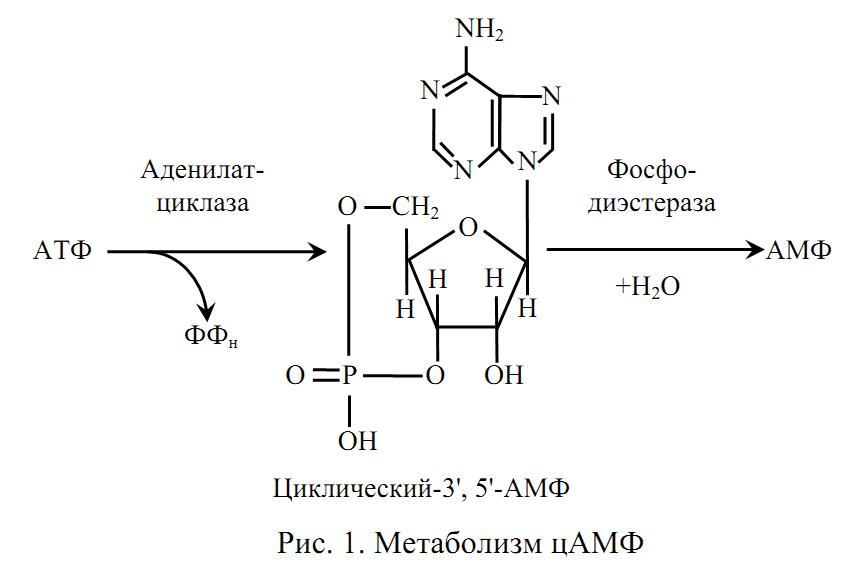
цАМФ (аденилатциклазный механизм)
цГМФ (гуанилатциклазный механизм)
Ионы Са2+
Инозитол-3-фосфат
ДАГ
NO
Иногда в клетке образуются и третичные посредники. Так, обычно ионы Ca2+ выступают в роли вторичного посредника, но при передаче сигнала с помощью инозитолтрифосфата (вторичный посредник) выделяющиеся при его участии из ЭПР ионы Ca2+ служат третичным посредником.
Свойства вторичных мессенджеров:
· Быстро образуются и выделяются, а так же быстро разрушаются
· Депонируются (ионы кальция)
· Продукция, выделение, обезвреживание находится под жёстким контролем внутриклеточных систем
Поверхностно-мембранный механизм действия гормонов (цАМФ)
 Образование
и инактивация цАМФ
Образование
и инактивация цАМФ
Поверхностно-мембранный механизм действия гормонов (ИФ3 и ДАГ)
 Цитозольный
механизм действия гормонов
Цитозольный
механизм действия гормонов