

Вопрос 36 Первичный транскрипт и его процессинг. Рибозимы как пример каталитической активности нуклеиновых кислот. Биороль.
Первичные транскрипты мРНК подвергаются ряду ковалентных модификаций до
их использования в качестве матрицы в синтезе белка.
1. Модификация 5'-конца: модификации пре-мРНК начинаются на стадии
элонгации. Когда длина первичного транскрипта достигает примерно 30 нуклеотидных
остатков, происходит кэпирование его 5'-конца.
Осуществляет кэпирование гуанилилтрансфераза, кот. гидролизует
макроэргическую связь в молекуле ГТФ и присоединяет нуклеотиддифосфатный остаток 5'- фосфатной группой к 5'-концу синтезированного фрагмента РНК с образованием
5',5'- фосфодиэфирной связи.
Последующее метилирование остатка гуанина в составе ГТФ с образованием
N7-метилгуанозина завершает формирование кэпа.
Значение: модифицированный 5'-конец обеспечивает инициацию трансляции
(рибосома для синтеза белка распознает только кэпированный конец), защищает от действия 5'-экзонуклеаз в цитоплазме, стимулирует сплайсинг (удаление интронов)
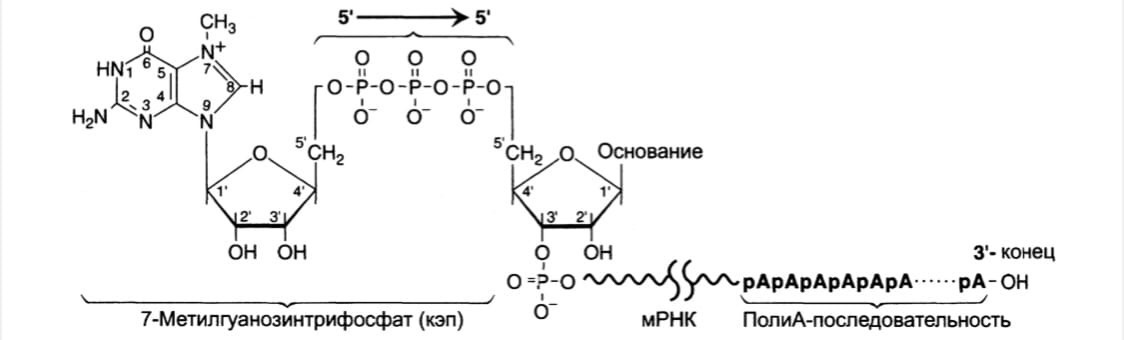
2. Модификация 3'-конца: 3'-конец большинства транскриптов, синтезированных
РНК-полимеразой II, также подвергается модификации, при которой специальным ферментом полиА-полимеразой формируется полиА-последовательность (полиА-«хвост»), состоящая из 100-200 остатков адениловой кислоты.
Сигналом к началу полиаденилирования является последовательность
-AAUAAA- на растущей цепи РНК.
Фермент полиА-полимераза, явл. экзонуклеазой,, разрывает 3'-фосфоэфирную связь после появления в цепи РНК специфической последовательности -AAUAAA
К 3'-концу в точке разрыва полиА-полимераза наращивает полиА-«хвост».
Наличие полиА-последовательности на 3'-конце облегчает выход мРНК из ядра и замедляет её гидролиз в цитоплазме.
Первичный транскрипт – строго комплементарная матрице нуклеиновая кислота
(пре-мРНК), содержащая экзоны и интроны.
Интроны – некодирующие последовательности нуклеотидов, присутствующие в ДНК, но не входящие в состав зрелой мРНК.
Экзоны – кодирующие последовательности, присутствующие в мРНК.
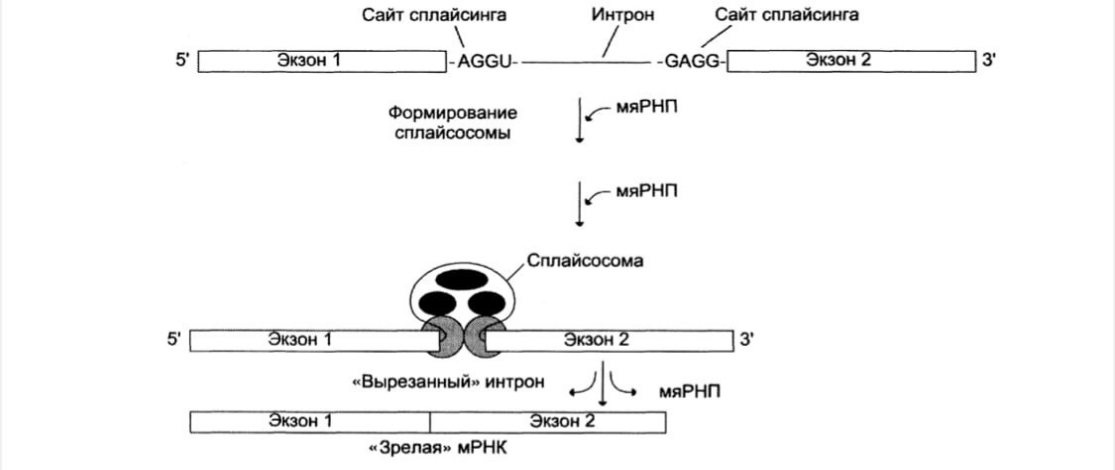
Сплайсинг – вырезание интронов из первичного транскрипта и сшивание концов
экзонов друг с другом. В результате сплайсинга, происходящего в ядре, из первичных транскриптов мРНК образуются молекулы «зрелой» мРНК, поступающие далее в цитоплазму к рибосомам для синтеза белка.
Процесс «вырезания» интронов протекает при участии малых ядерных
рибонуклеопротеинов (мяРНП), в состав кот. входит малая ядерная РНК (мяРНК), нуклеотидная цепь которой связана с белковым остовом, состоящим из нескольких протомеров.
На первой стадии процесса мяРНП связываются со специфическими
последовательностями первичного транскрипта (сайты сплайсинга), далее к ним присоединяются другие мяРНП.
Сплайсосома – десятки белков и 6 мяРНК, обеспечивающих сплайсинг
Она катализирует реакцию расщепления 3',5'-фосфодиэфирной связи на границе
экзона с интроном. Последовательность интрона удаляется, а два экзона соединяются.
Образование 3',5'-фосфодиэфирной связи между двумя экзонами катализируют мяРНК, входящие в структуру сплайсосомы.
Рибозимы – это каталитически активные молекулы РНК.
У прокариотов рибозимы осуществляют самосплайсинг, в процессе которого
интроны, расположенные между экзонами первичных транскриптов РНК «выщепляют сами себя» из молекулы РНК и сшивают кодирующие последовательности, образуя зрелую мРНК. Отличительной особенностью таких каталитически активных интронов считают высокую специфичность в отношении фосфодиэфирных связей определенных участков на РНК.
В клетках эукариот сплайсинг незрелых молекул мРНК осуществляет сплайсосома
с входящими в состав мяРНК.
В медицине рибозимы используют для расщепления специфических мРНК-
мишеней, что приводит к нарушению биосинтеза соответствующего белка.