

Вопрос 34 Повреждения и репарация днк. Виды повреждений. Способы репарации. Дефекты репарационных систем и наследственные болезни.
Репарация – процесс, позволяющий живым организмам восстанавливать
повреждения, возникающие в ДНК.
Репарационные механизмы основаны на том, что ДНК – двухцепочечная
молекула, т. е. в клетке есть 2 копии генетической информации.
Если нуклеотидная последовательность одной из двух цепей оказывается
повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена.
Если происходят повреждения, затрагивающие обе цепи ДНК,
то репарация не происходит.
Повреждения ДНК и их причины:
Ионизирующая радиация;
УФ-излучение;
Активные формы кислорода;
Химические агенты;
Спонтанные изменения.
Этапы репарации:
Выявление нарушения комплементарности цепей ДНК;
Устранение некомплементарного нуклеотида или только основания;
Восстановление целостности цепи по принципу комплементарности.
Виды репарации:
Прямая – обеспечивает прямое восстановление исходной структуры ДНК или удаление повреждения.
Например: удаление пиримидиновых димеров под действием фотолиазы.
Эксцизионная – происходит в несколько этапов сопровождается вырезанием большого фрагмента цепи ДНК.
Например: удаление продуктов реакций дезаминирования.
Виды повреждений:
1. Спонтанные – без участия каких-либо повреждающих факторов.
1.1 Ошибки репликации: происходят ошибки спаривания, и тогда вместо пары
нуклеотидов A–T, G–С в дочернюю цепь ДНК включаются некомплементарные нуклеотиды. ДНК-полимеразы δ, ε способны после присоединения очередного нуклеотида в растущую цепь ДНК делать шаг назад и вырезать последний нуклеотид, если он некомплементарен нуклеотиду в матричной цепи ДНК.
Пока основания нуклеотидных остатков в дочерней цепи неметилированы,
ферменты должны успеть выявить ошибку репликации и устранить её.
1 Этап: распознавание и удаление некомплементарного нуклеотида.
Происходит при участии специальных белков mut S, mut L, mut H.
Mut S находит неправильную пару и связывается с этим фрагментом.
Mut Н присоединяется к метилированному (по аденину) участку -GATC-,
расположенному вблизи некомплементарной пары.
Связующим между mut S и mut Н служит белок mut L, его присоединение завершает образование активного фермента.
Формирование комплекса mut S, mut L, mut H на участке, содержащем ошибку,
способствует проявлению у белка mut Н эндонуклеазной активности. Ферментативный комплекс гидролизует фосфоэфирную связь в неметилированной цепи
2 Этап: к свободным концам цепи присоединяется экзонуклеаза
Отщепляя по одному нуклеотиду в направлении от 3'- к 5'- концу дочерней цепи, она устраняет участок, содержащий некомплементарную пару.
3 Этап: брешь застраивает днк-полимераза β
4 этап: соединение основного и вновь синтезированного участков цепи
катализирует фермент ДНК-лигаза. Для успешного функционирования экзонуклеазы, ДНК-полимеразы β и ДНК-лигазы необходимо участие в репарации хеликазы и SSB-белков.
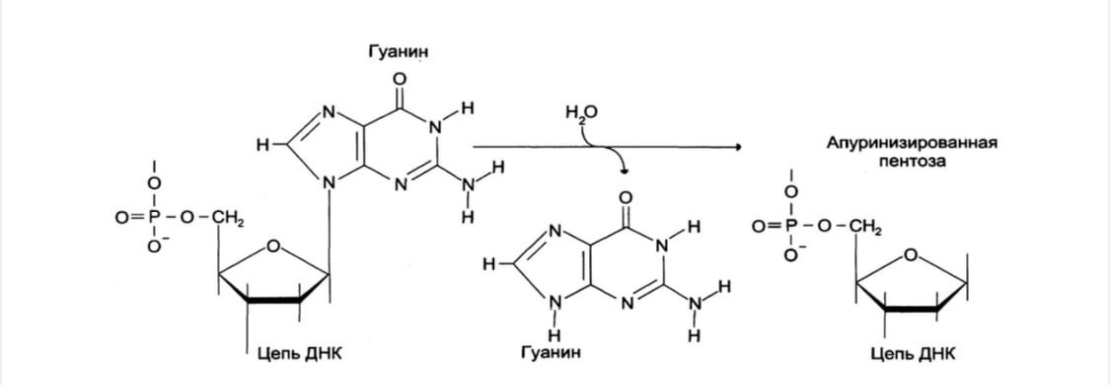
1.2 Депуринизация: потеря пуриновых остатков вследствие разрыва N-
гликозидной связи между пурином и дезоксирибозой. Образуется участок, лишённый азотистых оснований, названный АП-сайтом (апуриновый сайт). Этот тип повреждений устраняет фермент ДНК-инсертаза, который присоединяет к дезоксирибозе основание в соответствии с правилом комплементарности.
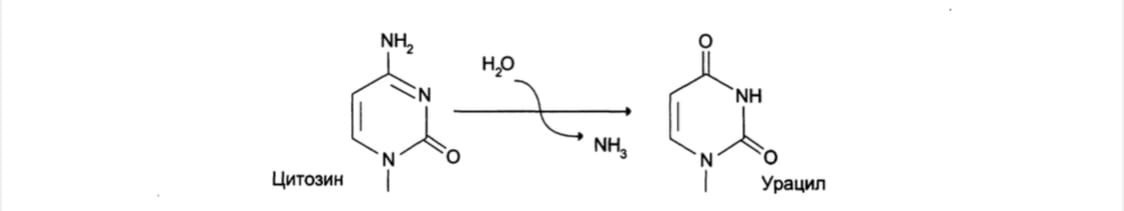
1.3 Дезаминирование: реакции дезаминирования цитозина и превращение его в
урацил, аденина в гипоксантин, гуанина в ксантин. Все продукты дезаминирования легко распознаются ферментами репарации
1 этап: ДНК-N-гликозилаза гидролизует связи между аномальным основанием и
дезоксирибозой, в результате образуется АП-сайт
2 этап: АП-сайт распознаёт фермент АП-эндонуклеаза
3 этап: как только в цепи ДНК возникает разрыв, АП-экзонуклеаза отщепляет от
цепи дезоксирибозу, лишённую основания
4 этап: в цепи ДНК появляется брешь размером в один нуклеотид и
ДНК-полимераза β к 3'-концу разорванной цепи присоединяет нуклеотид по принципу комплементарности
5 этап: ДНК–лигаза соединяет два свободных конца
2. Индуцируемые повреждения – в результате воздействия разнообразных мутагенных факторов как радиационной, так и химической природы.
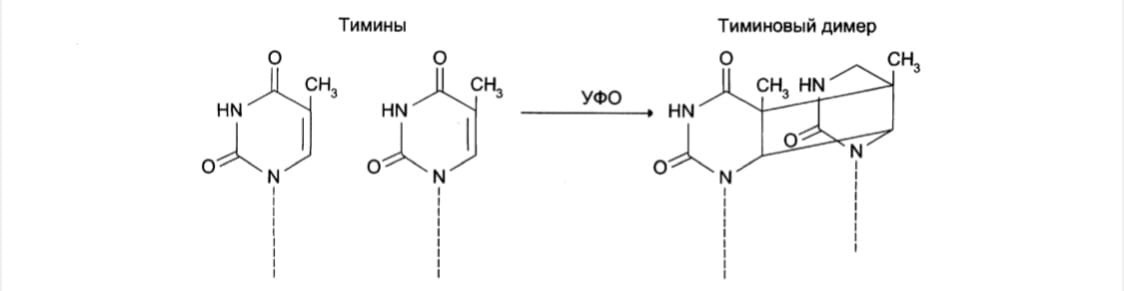
2.1 Образование димеров пиримидиновых оснований: под действием УФО
двойная связь между С5 и С6 атомами углерода в составе пиримидиновых оснований (тимине и цитозине) может разрываться. Атомы углерода остаются связанными одной связью. Расстояние между параллельными плоскостями оснований полинуклеотидной цепи, в которых произошёл разрыв – формируется циклобутановое кольцо.
Удаление пиримидиновых димеров происходит под действием фотолиазы.
Фермент расщепляет вновь образовавшиеся связи между соседними пиримидиновыми основаниями и восстанавливает нативную структуру.
2.2 Повреждения оснований ДНК химическими мутагенами: алкилированию,
окислению, восстановлению или связыванию основания с формамидными группировками.
1 этап: ДНК-N-гликозилаза присоединяется к повреждённому основанию.
Ферменты гидролитически расщепляют N-гликозидную связь между изменённым основанием и дезоксирибозой, это приводит к образованию АП-сайта в цепи ДНК
2 этап: репарация АП-сайта может происходить или только при участии
ДНК-инсертазы, которая присоединяет к дезоксирибозе основание в соответствии с правилом комплементарности, или при участии всего комплекса ферментов, участвующих в репарации: АП-эндонуклеазы, АП-экзонуклеазы, ДНК-полимеразы β и ДНК-лигазы.
Дефекты репарационных систем и наследственные болезни:
Пигментная ксеродерма: нарушение репарации УФ-повреждений (снижена
активность ферментов, ответственных за удаление неправильных оснований, «застройку» бреши). Дефект репарационной системы проявляется в сверхчувствительности к УФ-свету, что приводит к появлению красных пятен на коже, переходящих в незаживающие коросты и нередко в рак кожи.
Трихотиодистрофия: повышенная фоточувствительность ДНК, вызванная
снижением активности фермента, участвующего в удалении димеров тимина. Симптомы заболевания: ломкость волос вследствие нехватки серы в белках волос и их луковиц; часто умственная и физическая отсталость; аномалии кожи и зубов.
Синдром Блума (глубокие поражения капилляров лица) – мутация ДНК-лигазы