

Вопрос 33 Элонгация и терминация репликации. Ферменты. Асимметричный синтез днк. Фрагменты Оказаки. Роль днк-лигазы в формировании непрерывной и отстающей цепи.
Элонгация и терминация осуществляется ДНК-зависимыми ДНК-полимеразами.
Матрица – цепи ДНК.
Субстраты и источники энергии для синтеза – нуклеотидтрифосфаты: дАТФ, дГТФ, дЦТФ и дТТФ, для активации которых необходим Mg2+
Синтез цепей ДНК происходит в направлении 5'→3' растущей цепи.
Продукт – дочерние цепи ДНК.
ЭЛОНГАЦИЯ (синтез)
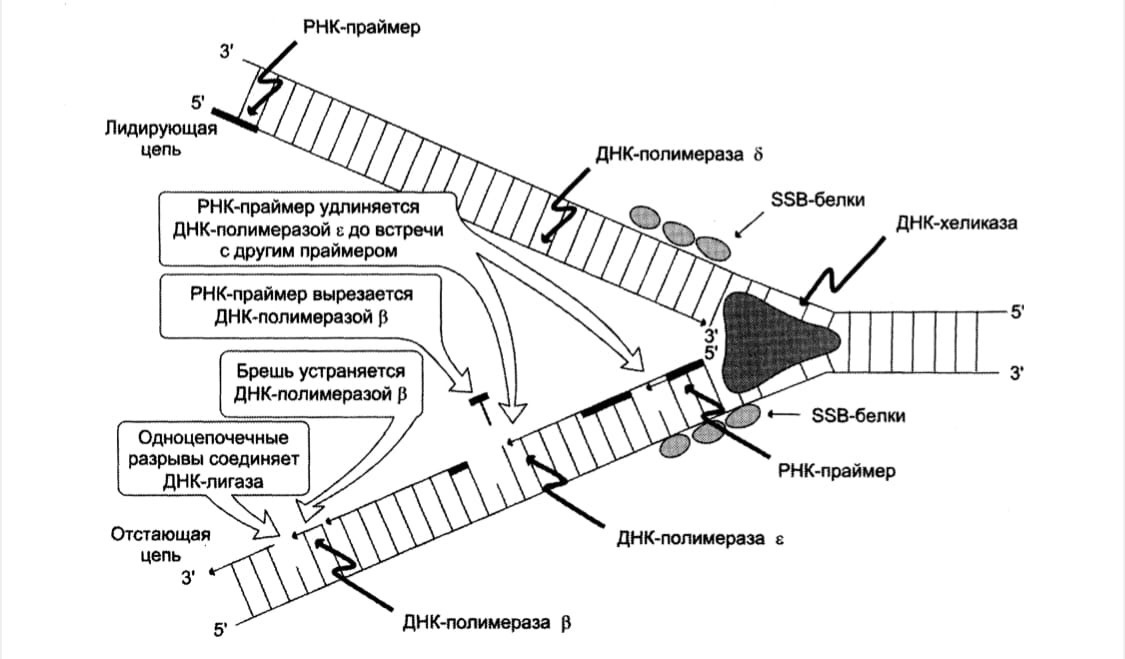
В синтезе эукариотических ДНК принимают участие 5 ДНК-полимераз (α,β,γ,δ,ε)
ДНК-полимеразы α, β, δ, ε участвуют в синтезе ДНК в ядре клеток.
ДНК-полимераза γ участвует в репликации митохондриальной ДНК.
ДНК-полимераза α инициирует репликацию (синтез праймера) – на этапе инициации
1. ДНК-полимераза δ последовательно наращивает цепь, присоединяя
соответствующие дезоксинуклеотиды (перемещается вдоль открытой нити ДНК, присоединяя вновь прибывшие нуклеотиды к свободному 3`ОН-концу новой нити ДНК, комплементарной матричной цепи);
Включение дезоксирибонуклеозидмонофосфатов сопровождается гидролизом
макроэрг.связей соответствующих нуклеозидтрифосфатов и отщеплением пирофосфата. Эта энергия используется на образование 3’-5’-фосфодиэфирной связи между последним нуклеотидом растущей цепи и присоединяемым
Матричная цепь считывается в направлении 3’→5’, а дочерняя синтезируется в
направлении 5’→ 3’ (антипараллельность)
2. Асимметричный синтез днк в направлении 5’→ 3’
В каждой репликативной вилке идёт одновременно синтез двух новых (дочерних)
цепей. Ферментативный комплекс ДНК-полимераз функционирует так, что одна из цепей растет с опережением относительно другой.
Лидирующая цепь - образуется в виде непрерывного длинного фрагмента, т.к её
направление синтеза цепи ДНК совпадает с направлением движ-я репликативной вилки.
Запаздывающая цепь – образуется в виде серии коротких фрагментов – «
фрагменты Оказаки». На ней синтез идёт в направлении 5'→3', но против движения репликативной вилки (ведется ДНК-полимеразой ε)
ТЕРМИНАЦИЯ – происходит при встрече 2х репликативных вилок
1. ДНК-полимераза β удаляет праймеры, постепенно
2. Застраивание брешей, образовавшихся после вырезания праймеров
дезоксирибонуклеотидами: на лидирующей цепи 1 праймер – присоединяет к ОН-гр на 3’-конце нуклеотиды в кол-ве, равному, вырезанному праймеру, на отстающей – праймер на каждом фрагменте Оказаки (фермент ДНК-полимераза β);
3. Сшивание фрагментов ДНК (фермент ДНК-лигаза).
Фермент ДНК-лигаза катализирует образование фосфодиэфирной связи между
3'-ОН-группой дезоксирибозы одного фрагмента цепи ДНК и 5'-фосфатом следующего фрагмента. Реакция протекает с затратой энергии АТФ. Таким образом, из множества фрагментов Оказаки образуется непрерывная цепь ДНК.
Метилирование ДНК происходит после завершения репликации. Метилируются
нуклеотиды в определенных последовательностях (например аденин всегда метилируется в последовательностях -GATC-). Метилирование не нарушает комплементарности цепей.
Наличие метильных групп в ДНК необходимо для формирования стр-ры хромосом,
а также для регуляции транскрипции генов. В течение некот. времени метилирование по Аденину происходит только в матричной цепи, это различие используется ферментами репарации для исправления ошибок в синтезированной цепи.