

Вопрос 35 Транскрипция. Характеристика компонентов системы синтеза рнк. Структура днк-зависимой рнк-полимеразы: роль субъединиц (2ββ′). Инициация процесса. Элонгация, терминация транскрипции.
Транскрипция – первая стадия реализации генетической информации в клетке.
Это синтез всех видов РНК на основе матрицы ДНК
Транскрипция у эукариот происходит в ядре.
Принципы транскрипции:
Комплементарность – РНК-полимераза синтезирует кмоплементарную реплику с
транскрибируемого участка ДНК (G ≡ C, A=U и Т=А).
Беззатравочность – транскрипция начинается с НТФ и не требует
затравочных олигонуклеотидов
Синтез цепи РНК идёт от 5'- к З'-концу, при этом матричная цепь ДНК всегда
антипараллельна синтезируемой нуклеиновой кислоте.
Ассиметричность – транскрибируется лишь одна из цепей ДНК (матричная цепь)
(Вторая, комплементарная ей цепь – кодирующая)
Рибонуклеозидтрифосфаты (ЦТФ, ГТФ, АТФ, УТФ) – субстраты и источники
энергии, необходимые для протекания полимеразной реакции, образования 3',5'-фосфодиэфирной связи между рибонуклеозидмонофосфатами.
Промоторы – определённые последовательности (сайты) ДНК, с которых
начинается синтез молекул РНК.
Терминаторы – терминирующие участки, в которых завершается синтез РНК.
Транскриптон – участок ДНК, ограниченный промотором и сайтом терминации,
представляет собой единицу транскрипции. В каждом транскриптоне присутствует неинформативная зона, которая содержит специфические последовательности нуклеотидов, с которыми взаимодействуют регуляторные транскрипционные факторы.
Транскрипционые факторы – белки, взаимодействующие с регуляторными
сайтами и ускоряющие или замедляющие процесс транскрипции.
Соотношение информативной и неинформативной частей в транскриптонах
У эукариот составляет в среднем 1:9
У прокариотов 9:1
Соседние транскриптоны могут быть отделены друг от друга
нетранскрибируемыми участками ДНК. Разделение ДНК на множество транскриптонов позволяет осуществлять с разной активностью транскрипцию разных генов.
Биосинтез РНК осуществляется ДНК-зависимыми РНК-полимеразами.
РНК-полимераза I, синтезирующая пре-рРНК;
РНК-полимераза II, ответственная за синтез пре-мРНК;
РНК-полимераза III, синтезирующая пре-тРНК.
РНК-полимеразы – олигомерные ферменты, состоящие из 5 субъединиц– 2 α,β,β',σ.
Это holo-фермент (полный фермент). Без -фактора (2 α, β, β') – core-фермент.
Стадии узнавания и связывания, а также инициации осуществляются holo-ферментом.
Элонгация и терминация осуществляются core-ферментом.
Роль субъединиц: две α-субъединицы – каркас РНК-полимеразы, к ним крепятся
остальные субъединицы; β' -субъединица отвечает за прочное связывание с ДНК за счет кластера положительно заряженных аминок-т. В β-субъединице находятся два каталитических центра. Один отвечает за инициацию, а другой – за элонгацию.
Субъединица σ «сигма» выполняет регуляторную функцию, это один из факторов
инициации транскрипции. РНК-полимеразы I, II, III, узнающие разные промоторы, содержат разные по строению субъединицы σ.
ИНИЦИАЦИЯ ТРАНСЛЯЦИИ
1. Активация промотора происходит с помощью большого белка – ТАТА-фактора,
взаимодействующего со специфической последовательностью нуклеотидов, входящего в состав промотора – ТАТА-бокс
2. Присоединение ТАТА-фактора облегчает взаимодействие промотора с
РНК-полимеразой. Факторы инициации вызывают изменение конформации
РНК-полимеразы и обеспечивают раскручивание примерно одного витка спирали ДНК, т.е. образуется транскрипционная вилка, в которой матрица доступна для инициации синтеза цепи РНК.
3. После того как синтезирован олигонуклеотид из 8-10 нуклеотидных остатков,
σ-субъединица отделяется от РНК-полимеразы, а вместо неё к молекуле фермента присоединяются несколько факторов элонгации.
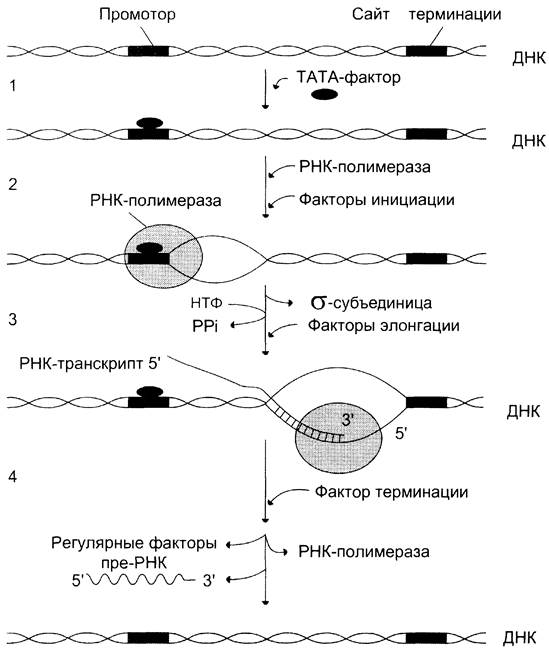
1 – присоединение ТАТА-фактора к промотору.
2 – образование транскрипционной вилки;
3 – элонгация; 4 – терминация
ЭЛОНГАЦИЯ
Факторы элонгации повышают активность РНК-полимеразы и облегчают
расхождение цепей ДНК. Синтез молекулы РНК идёт от 5'- к З'-концу комплементарно матричной цепи ДНК. На стадии элонгации, в области транскрипционной вилки, одновременно разделены примерно 18 нуклеотидных пар ДНК.
Растущий конец цепи РНК образует временную гибридную спираль, около 12 пар
нуклеотидных остатков, с матричной цепью ДНК. По мере продвижения РНК-полимеразы по матрице в направлении от 3'- к 5'-концу впереди неё происходит расхождение, а позади – восстановление двойной спирали ДНК.
ТЕРМИНАЦИЯ
Раскручивание двойной спирали ДНК в области сайта терминации делает его
доступным для фактора терминации. Завершается синтез РНК в строго определенных участках матрицы – терминаторах (сайты терминации).
Фактор терминации облегчает отделение первичного транскрипта (пре-мРНК),
комплементарного матрице, и РНК-полимеразы от матрицы. РНК-полимераза может вступить в следующий цикл транскрипции после присоединения субъединицы σ.
У бактерий существует 2 механизма терминации транскрипции:
ρ-зависимый механизм, при котором ρ-белок дестабилизирует водородные связи между матрицей ДНК и мРНК, высвобождая молекулу РНК.
ρ-независимый, при котором транскрипция останавливается, когда только что синтезированная молекула РНК формирует стебель-петлю, за которой расположено несколько урацилов (....УУУУ), что приводит к отсоединению молекулы РНК от матрицы ДНК.