

The embryonic development in birds
Birds produce few, large, yolk-rich eggs with protective membranes and shells. Thus, the ova in birds are referred to as polylecithal, pronounced telolecithal ova.
Cleavage. During incomplete (meroblastic, partial), diskoidal cleavage of the telolecithal egg the yolk does not divide. Cytoplasmic divisions are limited to the animal pole of the zygote (Fig.16)/ A series of cleavage divisions result in appearance of blastomeres. Blastomeres are arranged in the form of a disk, which lies over the yolk mass. This blastula is termed as a diskoblastula. The roof of the diskoblastula is formed by the blastodisk, and the bottom is the superficial layer of non-dividing yolk, which is termed as periblast.
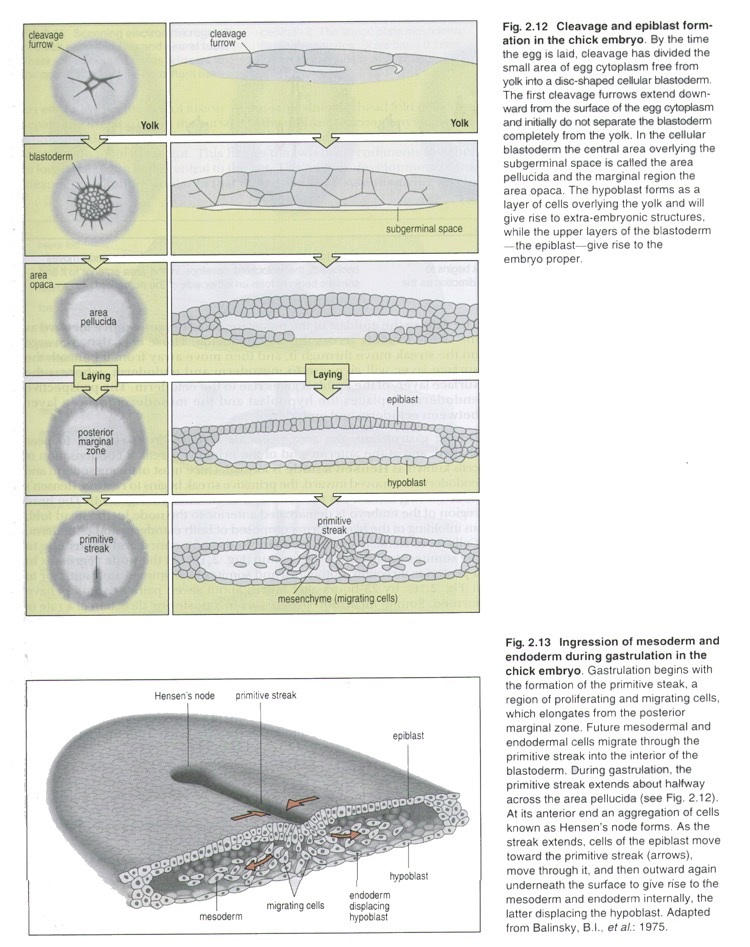 Fig.16.
Fig.16.
Gastrulation in birds proceeds by combination of delamination and immigration.
The first phase of gastrulation is delamination (Fig.17); it results in splitting off the internal layer – hypoblast (primitive endoderm), which will give rise to the extra-embryonic endoderm. At first the gut endoderm consists of round or irregular-shaped cells, which are arranged disorderly. Gradually the cells close up and form more or less regular single-layered plate. The rest of the germ disk forms epiblast (primitive ectoderm). Subsequently a cleft arises, which distinctly separates hypoblast from epiblast. The blastocoele is the thin cavity between the epiblast and hypoblast.
Further, cells of the central part of epiblast proliferate intensively, close up tightly and become more highly-prismatic in shape, forming the thickening of the central part of blastodisk. The epiblast contains the cell material of presumptive epidermal ectoderm, notochord, neural tube and mesoderm and endoderm of the embryo body. The thickened part of the blastodisk, consisting of two layers (the external one - epiblast, and the internal one - hypoblast), is termed as the germ disk. The germ disk is just the material, which forms the embryo body. The marginal parts of the blastodisk are extra-embryonic ecto- meso- and endoderm.

Fig.17. The diskoblastula in the chick (the vertical section).
1) The germ disk; 2) epiblast; 3) hypoblast; 4) yolk
The second phase of gastrulation in birds is immigration (Fig.18). It consists of complicated movements of cells of the epiblast. The cells start to move from the cranial (head) edge toward the caudal (tail) edge of the embryo. Two streams of moving cells come across one another at the median line on the caudal part of the embryo. Then the streams fuse, turn and move toward the cranial part along the median line of the body, forming thickened cell cord - primitive streak. The anterior broaden part of the primitive streak is termed as the Hensen' node (the primitive knot), which is effectively the equivalent of the dorsal lip of the amphibian blastopore.
A depression that forms within the streak is the primitive groove. The primitive groove is a dip through which the cells of epiblast move deep into and migrate on both sides of the primitive streak. Initially, the cells migrate anteriorly and ventrally forming the foregut endoderm, and displace the hypoblast cells. Gradually more epiblastic cells migrate on both sides of the primitive streak intervening between endoderm and ectoderm. These cells give rise to the middle germ layer - mesoderm.
 B
B
Fig.18. Gastrulation in Birds
A – formation of the primitive streak and the primitive node: 1) the primitive node; 2) the primitive pit; 3) the primitive streak; 4) the primitive groove
B – formation of mesoderm (immigration).
More epiblast cells ingress and form the head mesoderm and the anterior part of the notochord. The notochord is formed from some of the mesodermal cells in a middle layer between the ectoderm and the endoderm along the primitive streak. Following this, the primitive streak regress, moving Hensen’s node posteriorly. As it moves, the more posterior regions of the notochord develop. At its most posterior position, Hensen’s node forms the anal region. Notochord establishes the future axis of the vertebral column.
In the process of gastrulation, mesenchyme, embryonic connective tissue is forming. Mesenchymal cells are mainly mesodermal in origin but may be derived from the other germ layers. So, the cells migrate from their site of origin, surrounding and penetrating developing organs. Mesenchyme consists of loosely arranged stellate cells suspended in a gelatinous matrix. Their processes make contact with those of adjoining cells.
At the end we have three layers, the endoderm which has displaced the hypoblast, the ectoderm, which is what is left of the epiblast, and lots of mesoderm in between. Together the three germ layers and the mesenchyme give rise to all tissues, all organs and organ systems of the developing organism.
Thus, after gastrulation there follows a phase of histo- and organogenesis in which the primordial organs make their appearance. This phase gradually merges into the periods of growth and differentiation - both histological and functional.
At the beginning the formation of the axial primordial organs takes place. The complex of the axial organs includes: notochord, neural tube, mesodermal somites (paraxial mesoderm).
Neurulation (Fig.19) is the development of the neural tube form the neuroectoderm. Nerve tissue develops from embryonic ectoderm that is induced to differentiate in this direction by the underlying notochord. At first, a neural plate forms; then the edges of the plate thicken, forming the neural groove. The edges of the groove grow toward each other and ultimately fuse, forming the neural tube. This structure gives rise to the entire central nerve system. Some cells lateral to the neural groove, making up the neural crest, undergo extensive migrations and give rise to most of the peripheral nervous system, as well as a number of other structures (melanocytes, medulla of the adrenal gland, cells of the diffuse endocrine system. The rest of intra-embryonic ectoderm gives rise to the skin epidermis and its derivatives.
 Fig.19
Fig.19
The circumferential continuity of the endoderm is restored with the formation of a tube the definitive gut.
Alongside the neural tube and the notochord, mesoderm forms metamerically arranged somites (Fig.20). More laterally is a strip of intermediate mesoderm (syn.: nephrogonotome, or segment stalk). Pairs of somites and nephrotomes are metamerically segmented portions of the mesoderm. The splanchnotome (or lateral plate mesoderm) is a non-segmented portion of the mesoderm, placed the most laterally.
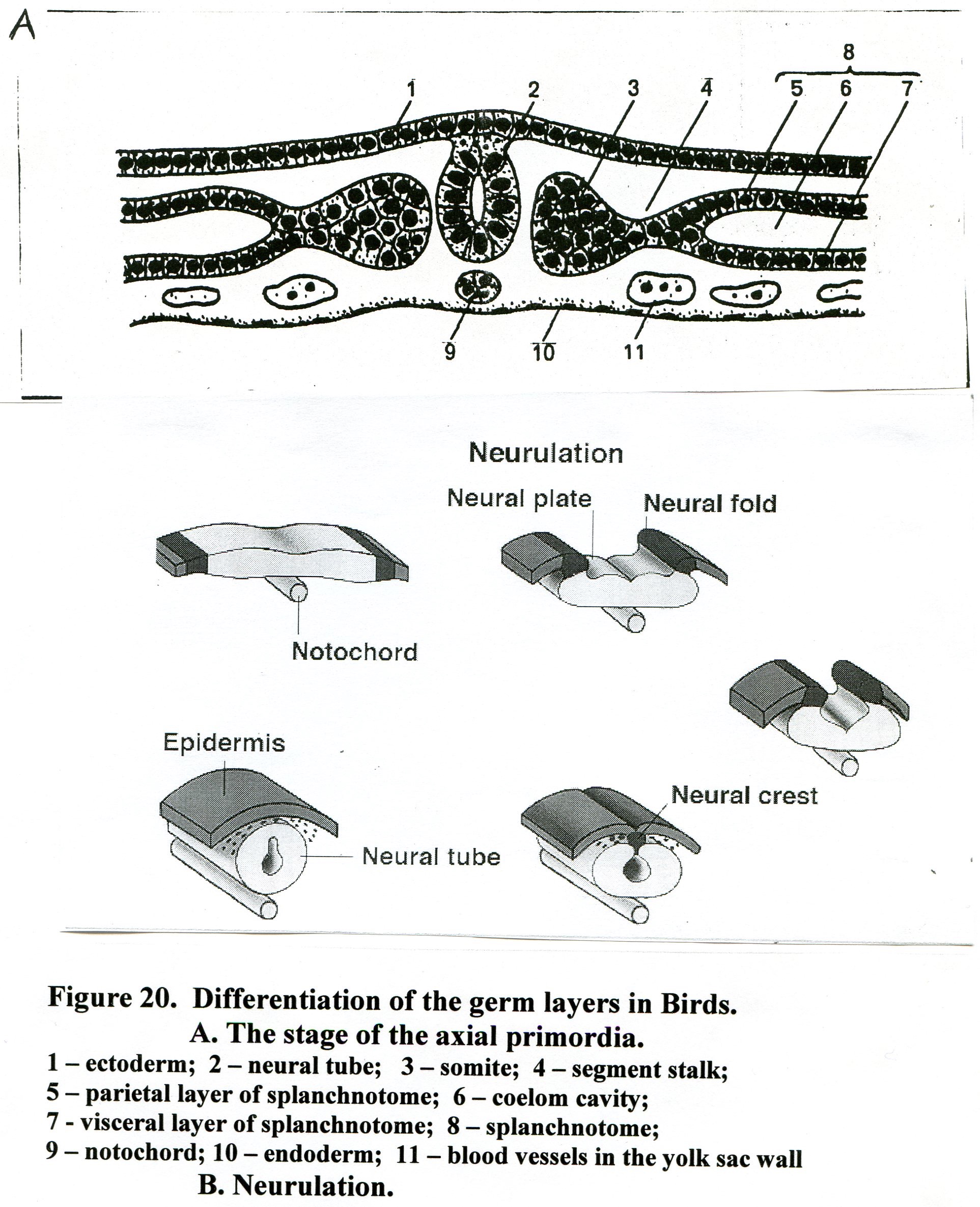
Fig.20. Differentiation of the germ layers in Birds
ectoderm; 2) nerve tube; 3) somite; 4) nephrogonotome (segment stalk); 5) parietal layer of splanchnotome; 6) coelomic cavity; 7) visceral layer of splanchnotome; 8) splanchnotome (lateral mesoderm); 9) notochord; 10) endoderm; 11) blood vessels in the yolk sac wall.
Further development involves subsequent differentiation of the mesoderm. The somite divides into dermatome, myotome and sclerotome (Fig.21). The dermatome, outer mass of cells, gives rise to the connective tissue layer of the skin (the dermis) and subcutaneous tissues. Cells proliferating from the dorsal part of the inner wall of the somite form the myotome and give rise to somatic muscle. The cells of sclerotome (the ventral part of the inner wall) migrate medially, envelop the notochord and contribute to the axial skeleton and meninges.
The segmented stalks will give rise to the kidneys and gonads.
Fig.21. . Differentiation of the germ layers in Birds
ectoderm; 2) nerve tube; 3) dermatome; 4) myotome; 5) sclerotome; 6) nephrogonotome;
7) notochord; 8) aorta; 9) coelom; 10) amniotic fold; 11) extraembryonic ectoderm;
12) parietal layer of splanchnotome; 13) body fold; 14 ) visceral layer of splanchnotome;
15) endoderm; 12) blood vessels in the yolk sac wall.
The splanchnotome splits onto parietal and visceral layers, which bound coelom cavities. The parietal layer of splanchnotome is in contact with ectoderm (somatopleure), and the visceral layer is in contact with endoderm (splanchnopleure). The lateral plate mesoderm gives rise to serous lining of cavities: pericardium, pleura, peritoneum, as well as cardiac muscle (myoepicardial plate, derived from the visceral layer of splanchnotome).
Mesenchyme cells are pluripotent, i.e. they may differentiate into a wide variety of cells. The mesenchyme gives rise to all types of connective tissue cells, as well as blood cells and blood vessels, hematopoietic organs, smooth muscle cells.
Endoderm gives rise to intestine epithelium and digestive glands (the liver, the pancreas), epithelial lining of the lungs, the urinary bladder etc.