

151. Рекции переаминирования и трансдезаминирования и их значение в метаболизме клеток.

Донорами аминогрупп явл. глутамат, аспартат, аланин. Путем трансаминирования происх.перераспределение аминного азота в тканях и органах. Наиб. Активно реакции идут в печени.Трансаминированию не подвергаются лизин, треонин и пролин.
Аминотрансферазы – АСТ – маркер инфаркта миокарда, АЛТ – маркер заболеваний печени.
Трансдезаминирование:
основной способ дезаминирования
большинства аминокислот. Идет в 2 стадии.

152.
Типы дезаминирования аминокислот и их
значение в клеточном обмене. Непрямое
дезаминирование, химизм процесса,
стадии, биологическое значение.
![]()
153. Химизм обезвреживания аммиака в печени. Биологическое значение этого процесса. (см.156в)
1 54.
Цикл мочевины.(см.156в) Связь орнитинового
цикла с превращениями фумаровой и
аспарагиновой кислот. Нарушения синтеза
и выведения мочевины. Гипераммониемии.
54.
Цикл мочевины.(см.156в) Связь орнитинового
цикла с превращениями фумаровой и
аспарагиновой кислот. Нарушения синтеза
и выведения мочевины. Гипераммониемии.
155. Трансаминирование, химизм процесса, специфичность аминотрансфераз. Диагностическое значение определения аминотрансфераз.
Донорами аминогрупп явл. глутамат, аспартат, аланин. Путем трансаминирования происх.перераспределение аминного азота в тканях и органах. Наиб. Активно реакции идут в печени.Трансаминированию не подвергаются лизин, треонин и пролин.
Аминотрансферазы – АСТ – маркер инфаркта миокарда, АЛТ – маркер заболеваний печени.
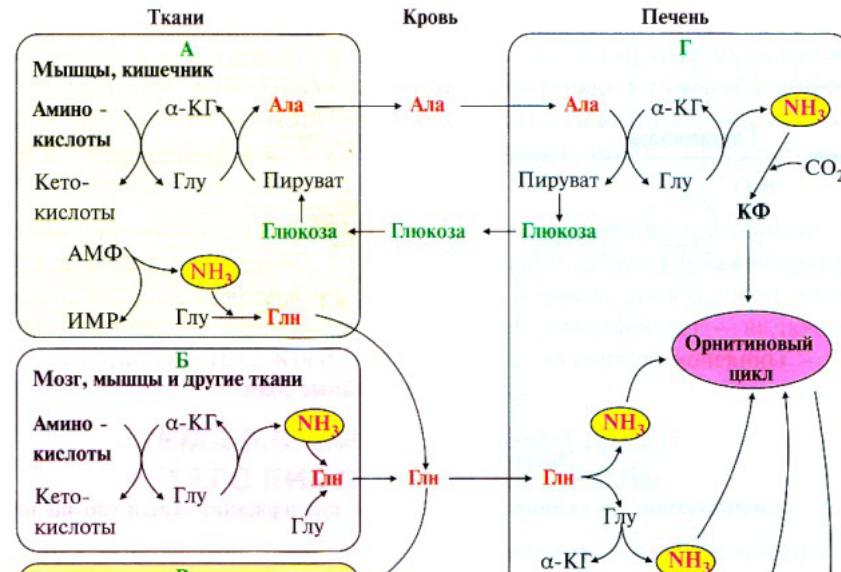
156.
Пути обезвреживания аммиака в организме.
Цикл мочевинообразования. Механизмы
обезвреживания аммиака в нервной и
мышечной ткани. Из
организма аммиак выводится в виде
конечных продуктов азотистого обмена:
мочевины, аммонийных солей. Основной
р-цией обезвреживания явл.синтез
глутамина.
![]()
Глутамин можно считать транспортной формой аммиака, он явл. нейтр.ам-к и способен легко проникать в кл.мембраны путем облегченной диффузии. В печени аммиак обезвреживается путем связывания с СЩ2 и образования карбамоилфосфата, используется 2АТФ. Затем карбамоилфосфат исп. В орнитиновом цикле.
Мочевая
кислота является фракцией остаточного
азота и конечным продуктом обмена
пуриновых оснований (аденина и гуанина).
М.к-та, выводимая с мочой отражает
поступление пуринов с пищей и распад
эндогенных пуриновых нуклеотидов.

Орнитиновый цикл в печени выполняет 2 ф-ции – превращение азота ам-к в мочевину, кот.экскретируется и помогает избежать накопления аммиака в организме.; синтез аргинина и пополнение его фонда в организме.
157. Глицин, его строение и роль в обмене веществ.
158. Основные пути метаболизма глицина. Глицин как важнейший донор углеродных фрагментов для биосинтезов.


159. Глутамат и аспартат, их химическое строение и роль в обмене веществ.
В организме исп.всеми клетками для синтеза пуриновых и пиримидиновых оснований. Амидные производные этих ам-к явл.транспортными формами аммиака из тканей в почки и печень. Кроме этого, глутаминовая к-та входит в состав глутатиона, ф-ции:перенос ам-к ч/з мембрану, медиатор. Асп.принимает участие в орнитиновом цикле, в р-циях трансаминирования и биосинтезе углеводов (гликогенная аминокислота)


160. Аспарагиновая и глутаминовая кислоты, основные пути метаболизма, биологическое значение глутатиона.


161. Строение аргинина и гистидина. Их роль в обмене веществ.
Аргинин. Ф-ции: -используется в синтезе креатина, который в виде креатинфосфата способен служить источником энергии для работы мышц человека и млекопитающих.
-служит источником NO в организме;
-служит
предшественником орнитина, из которого
синтезируются полиамины
спермидин и
спермин.
Аргинин
служит в организме источником оксида
азота (NO). Образование NO в клетках
катализирует сложный Са2+-зависимый
фермент NO-синтаза. В состав фермента
входит гем.




162. S-аденозилметионин, его химическое строение и роль в метаболизме клетки. Липотропные факторы(холин, метионин, витамин В12, фолиевая к-та) – в-ва, способствующие синтезу ФЛ и препятствующие синтезу ТАГ. При недостатке липотропных факторов развивается жировая инфильтрация печени.

SAM- активная форма метионина, присутствует во всех типах клеток. Участвуюет во многих р-циях метилирования, в частности в синтезе креатинина, карнитина, лецитина, адреналина, а так же в инактивации норм.метаболитов и обезвреживании токсич.в-в печени. В рез-те перемещения метильной группы и отщепления аденозина остается гомоцистеин.Метилирование с участием SAM под действием метилтрансфераз – один из способов инактивации биогенных аминов.

163.Роль цистеина и метионина в обмене веществ. Липотропные факторы (холин, метионин, витамин В12, фолиевая к-та) – в-ва, способствующие синтезу ФЛ и препятствующие синтезу ТАГ. При недостатке липотропных факторов развивается жировая инфильтрация печени.
Метионин-незаменимая
ам-к, необ.для синтеза белков, подвергается
транс- и дезаминированию. Особ.роль
метионина-метильная группа этой ам-к
исп.для синтеза целого ряда соединений
в р-циях трансметилирования, донором
метильной группы явл.SАM.

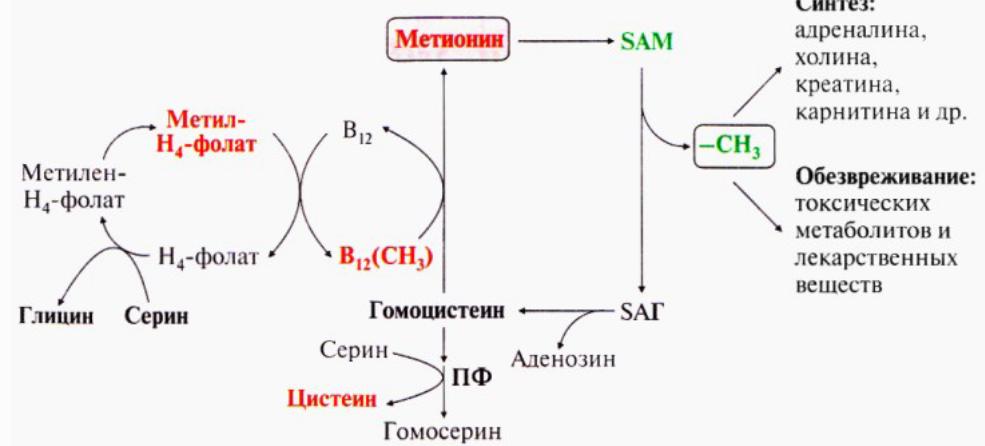
Метионин(донор серы) и серин необ.для синтеза цистеина. Обр.из гомоцистеина в ходе 2 реакций. Ген.дефект пиридоксальфосфата приводит к нарушению исп.гомоцистеина и превращ.в гомоцистин, который накапливаясь в кровь и тканях, выделяется с мочей вызывая гомоцистиинурию. Так же причиной заболевания мб гиповитаминох фолиевой к-ты или витаминов В6 и В12.