

39. Синтез насыщенных и ненасыщенных жирных кислот: локализации в клетке, необходимые источники и их происхождение, продукты реакции, участие витаминов. Регуляции синтеза жирных кислот гормонами
процесс активен первые 2ч после еды под действием гормона инсулина. Глю из крови →в клетки, где окисляется в результате образования ацетил-КоА в матриксе митохондрий, который должен включаться в биосинтез ЖК. Процесс синтеза ЖК осуществляется в ЦП, поэтому необходим перенос ацетил-КоА из митохондрий в ЦП: в матриксе митохондрий ацетил-КоА связывается с оксало-ацетатом → образуется цитрат, который легко пересекает митохондриальную мембрану и оказывается в ЦП, где распадается с высвобождением ацетил-КоА, который далее включается в синтез ЖК
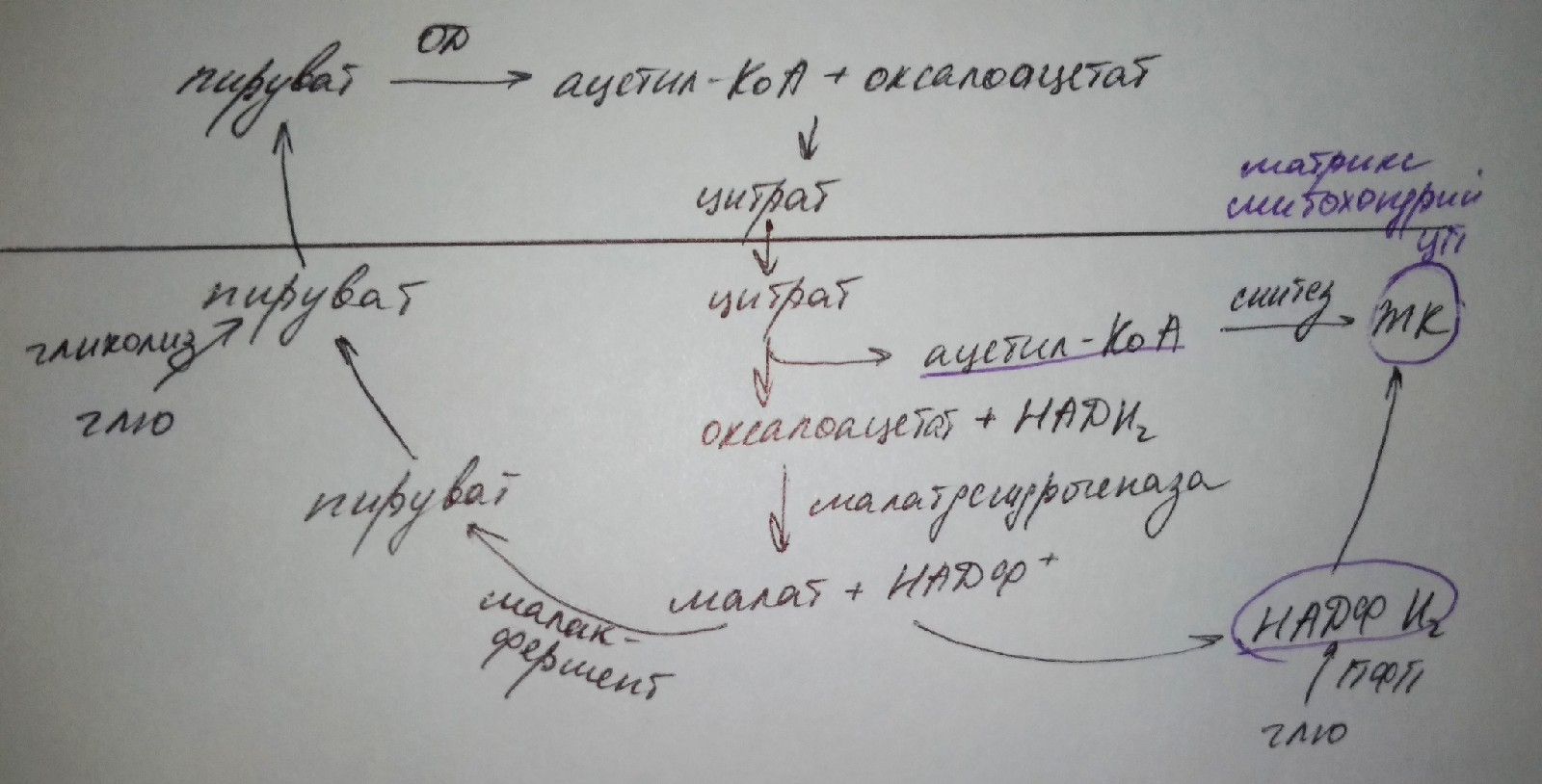
Таким образом для синтеза ЖК необходим
ацетил-КоА, НАДФН2, ферментный комплекс
полиметат-синтетаза, т. к. в клетках
человека преимущественно синтезируется
полиметиновая кислота. Для синтеза
необходимо осуществить несколько
циклов, в ходе каждоко уикла жирно-кислотная
цепь удлиняется на 2 углеродных атома,
источником которых является малонил-КоА.
В клетках мозга образуются еще более длинные ЖК, которые включаются в состав сфинго-фосфолипидов и гликолипидов.влияние инсулина
Активен после еды в ответ на гипергликемию, действует на скелетные мышцы, печень, жировая ткань.
В этих клетках активируются процессы:
1) гликолиз: глю окисляется пируват
2) окислительное декарбоксилирование пирувата до ацетил-КоА
3) пентозофосфатный путь окисления глю: образуется НАДФН2
4) синтез ЖК из ацетил-КоА при участии НАДФН2
5) гликолиз: глю окисляется фосфодиоксиацетон, который востанавливается до глицерол-3-ф
6) синтез ТАГиз глицерол-3-ф и ЖК
40. Синтез и использование организмом кетоновых тел в качестве источника энергии
В процессе ночного голода имеется дефицит глю, поэтому становится необходимым получение альтернативных источников энергии — кетоновых тел, которые окисляются клетками мозга, миокарда, ск. мышцами, почек. При этом большая часть глю сберегается преимущественно для эритроцитов и мб использована мозгом. В жировой ткани происходит распад ТАГ с образованием ЖК. ЖК → плазма и часть их поступает в клетки печени, где ЖК подвергается б-окислению с образованием ацетил-КоА, который в условиях дефицита год не может быть включен в цикл Кребса, т.к. оксало-ацетат, участвующий в глюконеогенезе и не может взаимодействовать с ацетил-КоА (1 реакция и ц.Кребса) поэтому ацетил-КоА используется на синтез кетоновых тел: ацетоацетата, б-гидроксибутерата и ацетона.
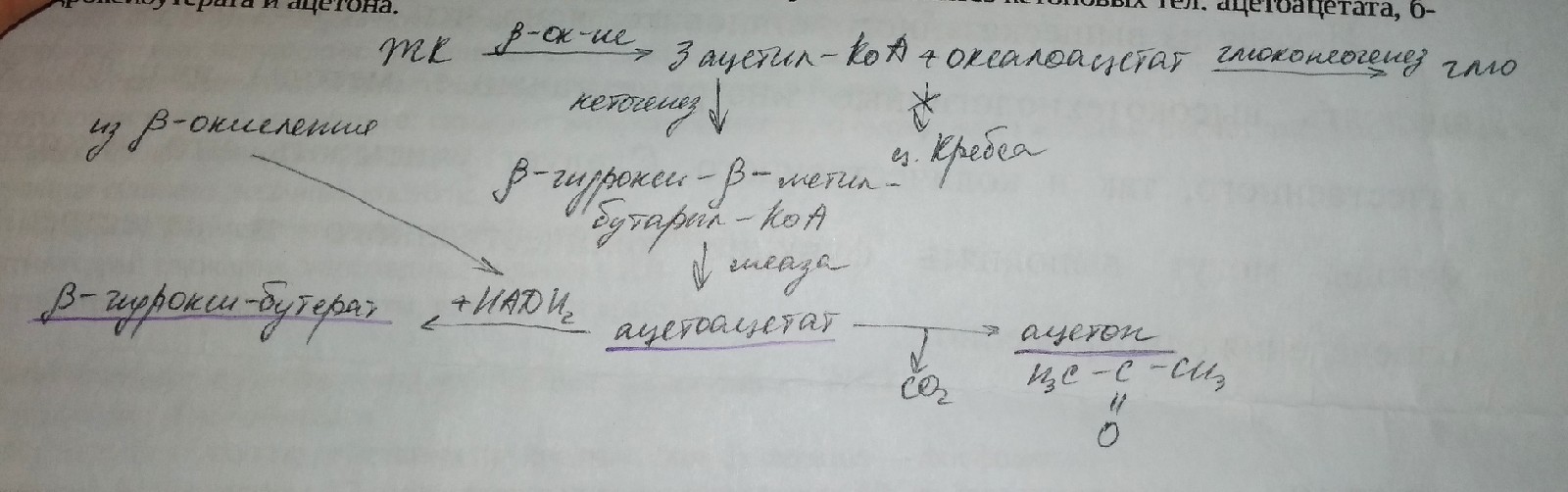

Процесс осуществляется в матриксе митохондриальных гепатоцитов, из кл. печени. Кетоновые тела выводятся в плазму → клетки ск.мышц, миокарда, почек, кишечника, мозга, где осуществляются следующие процессы
Клетки печени кетоновые тела не используют как источники энергии. Гепатоциты окисляют АК с целью энергообеспечения. При длительном голоде, длительной физической нагрузке, длительной лихорадке, сахарном диабете, токсикозе происходит ↓ использования глю, как источника энергии, поэтому при этих состояниях нарастает кетогенез Накопление кетоновых тел приводит к сдвигу рН в кислую сторону — кетоацидоз. Избыток кетоновых тел преимущественно удаляется в виде ацетона через кожу, лёгкие и через почки с мочой.