

активные и репрессивные модификации пространственно сегрегирован
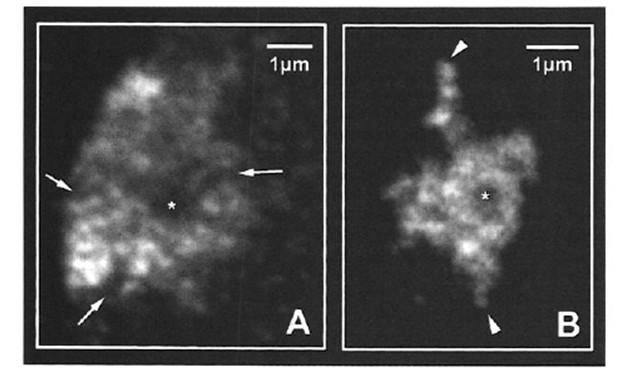
ользованием микроскопии высокого разрешения можно видеть, что хромосомные итории неоднородны. Они состоят из относительно компактных глобулярных домен ожно соответствующих ТАДам), разделенных пространством, в котором плотность атина существенно ниже).
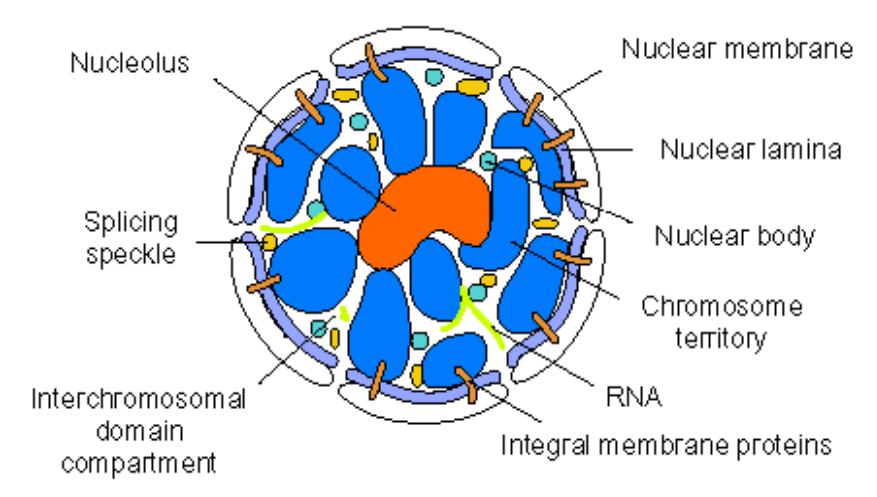
откуда берется ядерный матрикс?
Особенности позиционирования хромосомных территор и генов внутри хромосомных территорий
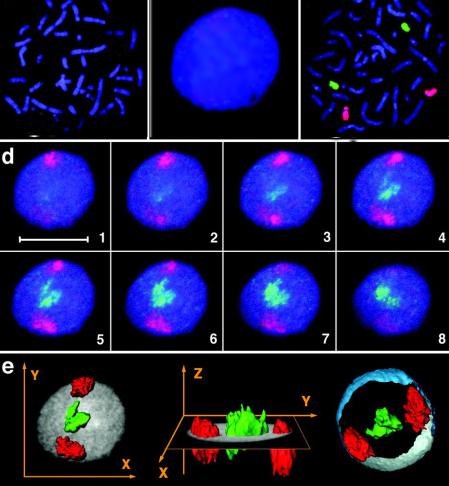
Разработка техники дифференциальной окраски хромосом и техники конфокальной микроскопии позволила анализировать взаимное пространственное расположение больших групп хромосом
Интерфазные хромосомы занимают в клеточном ядре относительно компактные неперекрывающиеся территории. Наиболее богатые активными генами
хромосомы располагаются ближе к центру ядра
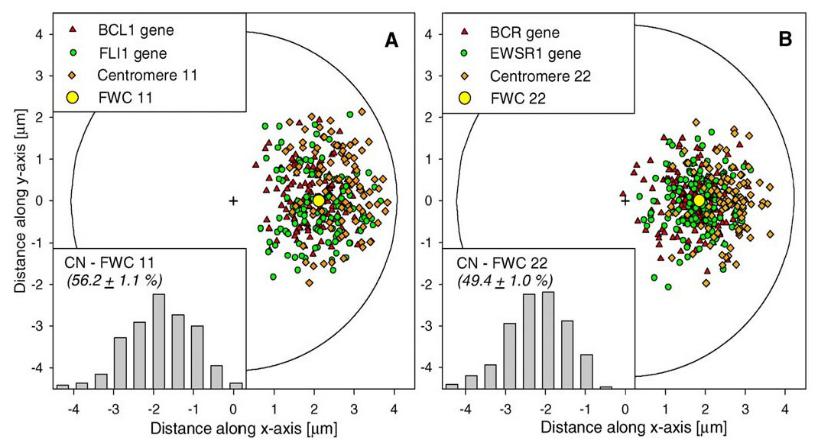
При оценке положения хромосмных территорий в ядре наиболее удобным параметром является расстояние между центром ядра и центром хромосомной территории (измеряется в % от радиуса ядра.
Хромосмома 22 находится ближе к центру ядра, чем хромосома 11. Внутри обоих хромосомных территорий центромеры располагаются на наибольшем удалении от центра ядра.
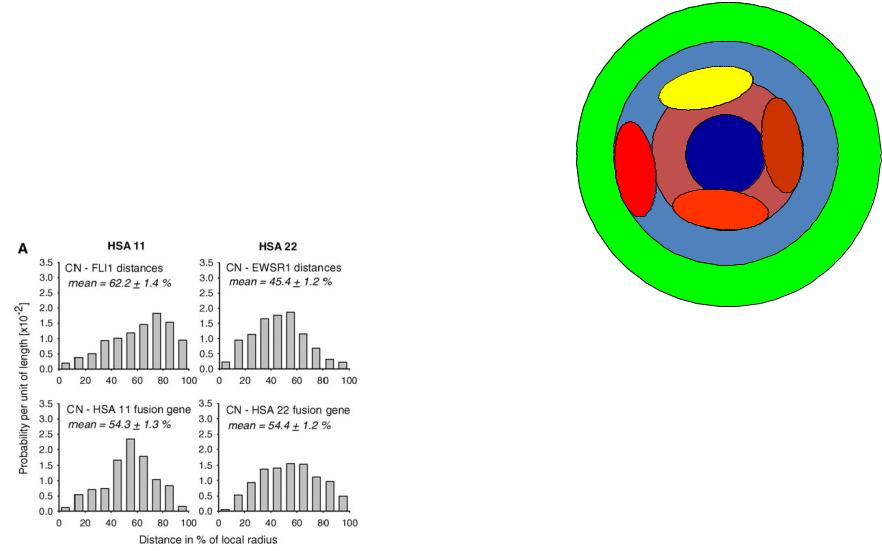
Внутри хромосомных территорий позиции генов не являются случайными
Соотносительные радиальные позиции генов FLI1 (HSA11) и EWSR1 (HSA22) различаются еще в большей степени, чем позиции соответствующих хромосомных территорий
Fused ген FLI1/EWSR1 занимает промежуточную радиальную позицию. Таким образом радиальная позиция хромосомной территории определяется совокупностью сигналов, распределенных по всей длине хромосомы
1 1
A S H
|
|
|
|
|
2 |
|
|
|
|
/2 |
|
|
|
|
1 |
|
|
|
|
1 |
|
|
|
|
A |
|
|
|
|
S |
|
|
|
|
|
H |
|
|
|
|
|
9 A S H
HSA22
Гены, находящиеся в одном и том же ядерном слое чаще учавствуют в реципроктных рекомбинациях,
чем гены, находящиеся в различных ядерных слоях.
Например, рекомбинации между ABL1 (HSA9) и BCR (HSA22) происходят существенно чаще, чем рекомбинации между FLI1 и EWSR1
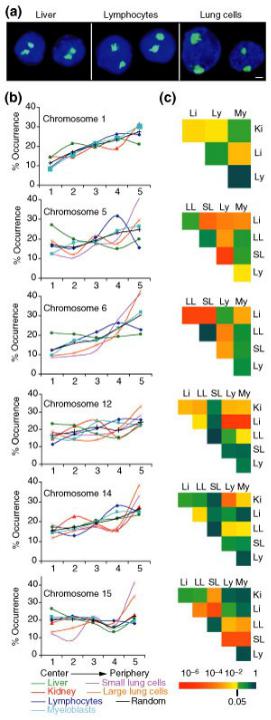
Предпочтительные радиальные позиции индивидуальных хромосом могут различаться в клетках, дифференцированных по разным путям
Хромосома 5 мыши занимает:
центральную позицию в клетках печени
промежуточную позицию в лимфоцитах
периферическую позицию в клетках легкого
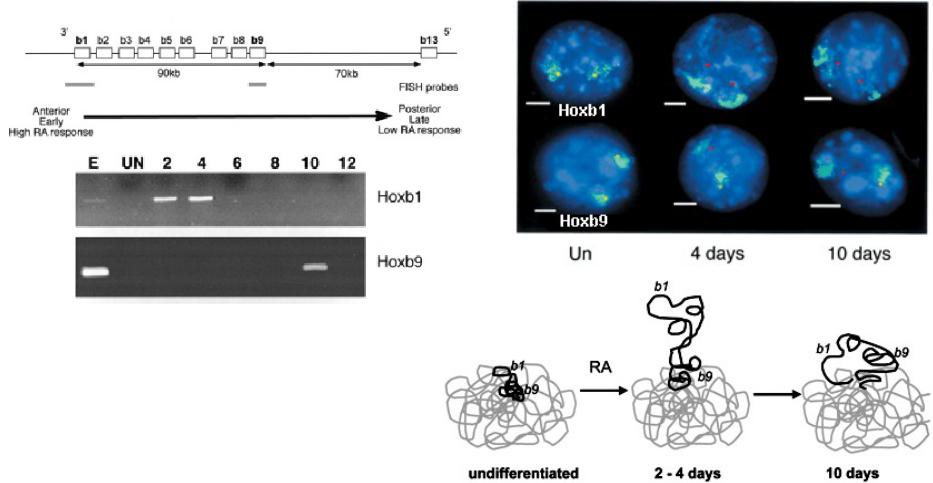
цы хромосомных территорий являются достаточно условными. В ряде случ аются вне хромосомных территорий
В ходе индуцированной ретиноевой кислотой дифференцировки мышиных ES клеток Hox гены транскрибируются в том порядке, в каком они расположены в геноме. При этом
происходит последовательное выпетливание из хромосомной территории соответствующих фрагментов ДНК