

Добавил:
Nemo_Nemorino
Опубликованный материал нарушает ваши авторские права? Сообщите нам.
Вуз:
Предмет:
Файл:Хроматин презентации / Lecture_2_2019.pptx
Привлечение к энхансеру гистондезацетилаз
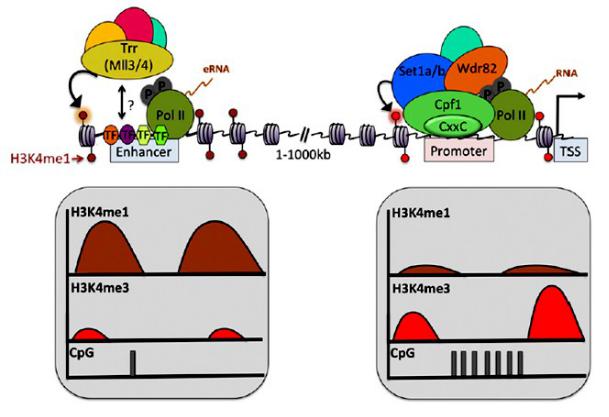
Почему на энхансерах нет триметилирования Н3К4, хотя энхансеры транскрибируютсмя Pol II?
Привлечение через посредство Pol II не является единственным механизмом привлечения Set метилаз
Set 1a/b в комплексе с CpF1 (узнает CpG-богатые последовательности) осуществляет преимущественно триметилирование Н3К4
MLL3/4 (Tritorax-like) осуществляет преимущественно монометилирование
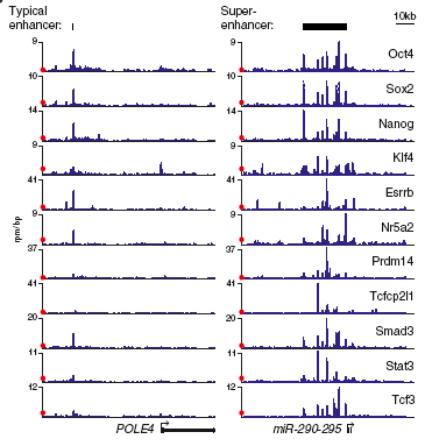
Супер-энхансеры
Блоки энхансеров, контролирующие работу генов, определяющих идентичность (тип дифференцировки) клеток. Содержат множественные места связывания мастер-регуляторов транскрипции
В мышиных стволовых клетках супер-энхансеры (всего 231) связывают Oct4, Sox2, Nanog и ряд других транскрипционных факторов, типичных для стволовых клеток