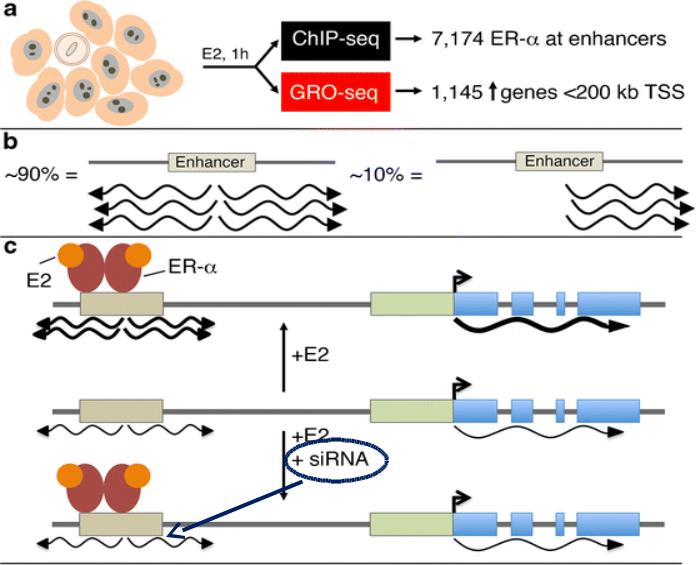
Позиции активных и потенциальных (репрессированных в данном типе клеток) энхансеров можно предсказать на основе анализа модификаций гистонов
Энхансер, активный в эмбриональных стволовых клетках
p300
H3K4me1
H3K27acet
работающие
энхансеры
неработающие
энхансеры
Энхансер, неактивный в эмбриональных стволовых клетках, но начинающий работать на более поздних стадиях развития
В ESC
p300
H3K4met1
H3K27me3
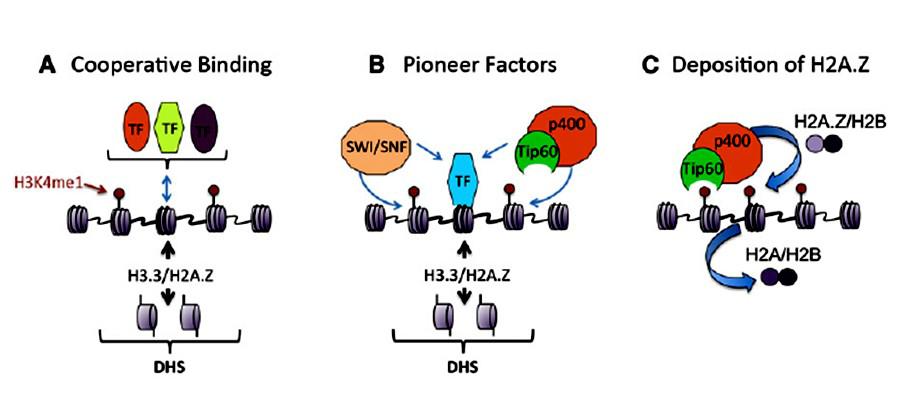
Создание особого хроматинового ландшафта на энхансерах может обеспечиваться рядом механизмов
1.Кооперативное связывание ряда транскрипционных факторов
2.Связывание транскрипционных факторов, которые узнают мишени на нуклеосомной ДНК
3.Привлечение коактиваторов, которые модифицируют гистоны и депонируют H3.3 и H2AZ
Хромодомен Tip60 специфически узнает H3K4 met1. Комплекс Tip60-p400 обеспечивает посадку и ацетилирование H2AZ
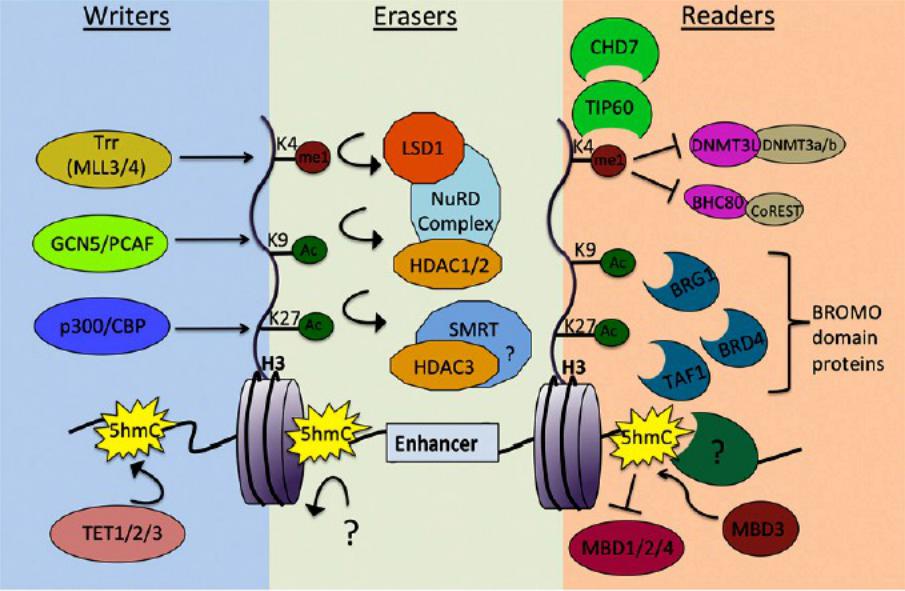
Ферменты, вносящие, удаляющие и считывающие модификации гистонов на энхансерах

Энхансеры служат местами посадки РНК полимеразы II, которая синтезирует различные типы некодирующей РНК.
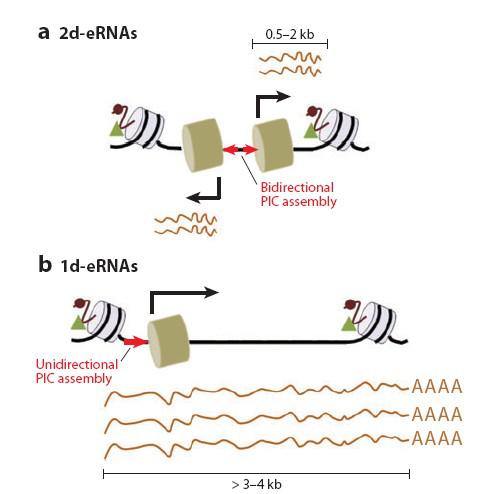
Существуют разные классы РНК, транскрибирующейся с энхансеров
короткая двунаправленная РНК (eRNA)
Длинная однонаправленная РНК полиаденилированная или не полиаденилированная сплайсированная или не сплайси-
рованная
Дискриминация от lincRNA представляется проблематичной

Наряду с короткими двунаправленными транскриптами (eRNA), внутригенные энхансеры могут направлять и снтез протяженных транскриптов в направлении транскрипции гена. Эти, укороченные транскрипты правильно сплайсируются и полиаденелируются, но, как правило, не транслируются
Molecular Cell (2012), doi:10.1016/j.molcel. 2011.12.021
meRNA
(multiexonic enhancer RNA)
R1-R4 – консервативные фрагменты, включающие участки гипечувствительности к ДНКазе I и эритроид-специфичные энхансеры. Ген Nprl3 не является эритроид-специфичным
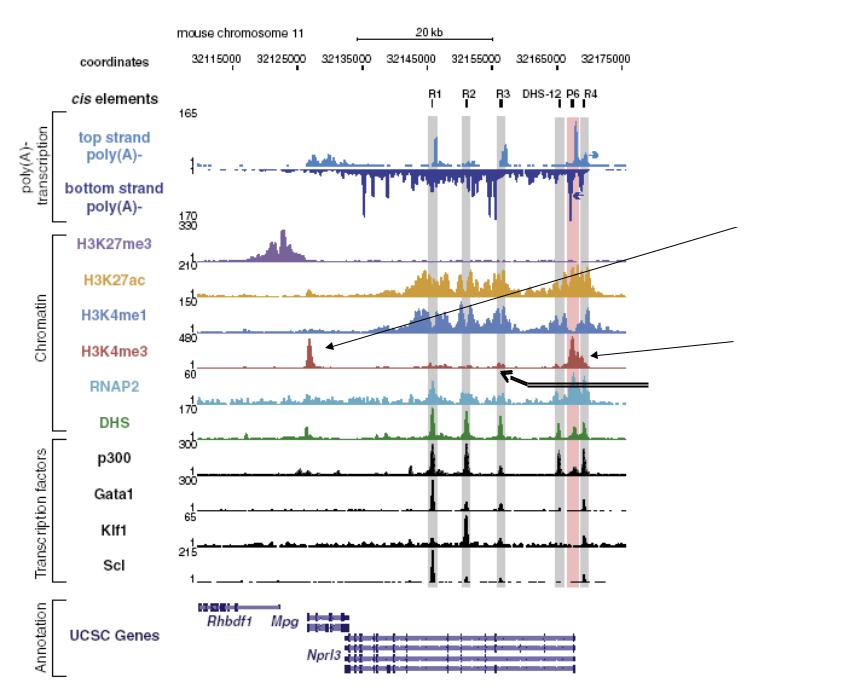
Промотор гена Mpg
Промотор гена Nprl3
Старт альтернативного транскрипта на R3 энхансере не содержит H3K4me3

Зачем нужна eRNA?
никакого функционального значения
важен процесс транскрипции, продукт не важен
важен продукт (eRNA)
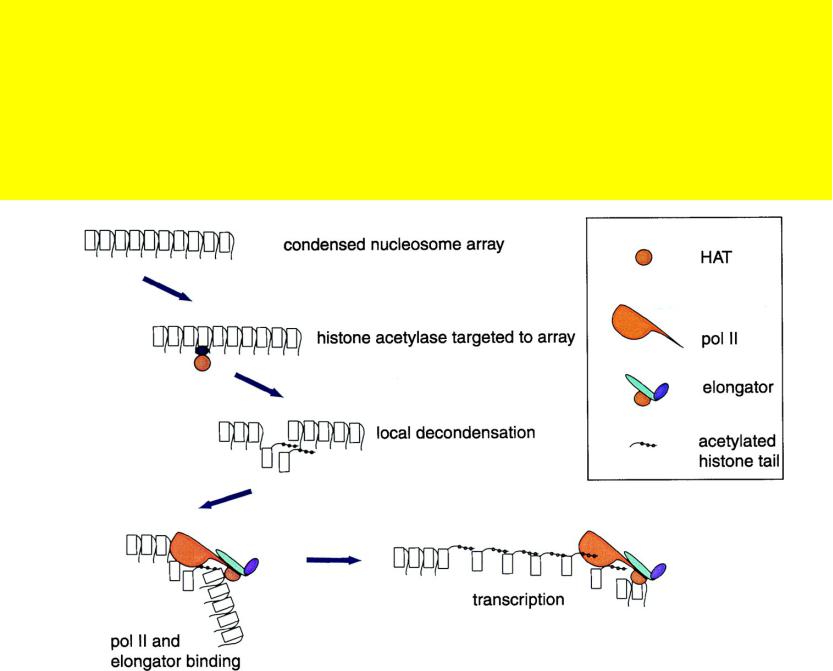
РНК-полимераза может служить “транспортным средством”, обеспечивающим
распространение активационных сигналов от LCR к промоторам индивидуальных генов
Travers, 1999