

Лекция 5
Репликация т-четных фагов
Общая структура: нам уже известна.
Геном: линейная двуцепочечная ДНК длиной 170000 п.н. с длинными прямыми концевыми повторами длиной около 3000 п.н. Могут быть кольцевые перестановки.
Мутации по 30 генам блокируют размножение вируса. Однако с результатами мутационного анализа надо быть осторожными, потому что продукты не всех генов непосредственно не участвуют в репликации ДНК, но мутации по ним блокируют размножение. Например, у фага φХ174 мутация по структурным белкам блокирует его размножение. К счастью, в случае Т-четных фагов все эти 30 генов участвуют в репликации.
Инициация репликации:
1.ori-зависимая
2.рекомбинационная
1.ori-зависимая инициация репликации.
Протекает не по обычной схеме. Обычно внутри генома вируса имеется выделенный элемент (ori), с которым связывается специальный белок (OBP), который способствует образованию праймосомы, а затем реплисомы. В случае Т-четных фагов в ДНК много ori. Карта Т-четных фагов кольцевая. Генетические расстояния определяются по частоте рекомбинаций между генетическими маркерами. Из-за пермутаций участки, удаленные в одной молекуле могут оказаться вблизи в другой, это легче объяснить, если представить ДНК в виде кольца.






C
D E
B
A












О


G F



Положение ori соответствует промоторам средних генов (конечно, промоторы не всех средних генов содержат ori). Если ori находится в районе промотора, то, видимо, инициация репликации происходит с участием РНК-полимеразы. Инициация рифампицин-чувствительная. Участвует клеточный фермент, своей у вируса нет. Вирусный белок Mot (Modification of transcription) способствует узнаванию ori-содержащих промоторов генов. Полимераза синтезирует участки короткой РНК, которые далее будут использованы как затравки при синтезе ДНК. РНК, синтезированная в других местах затравкой не служит. Только промоторного участка не достаточно для синтеза РНК, вблизи находится легкоплавкий участок ДНК, называемый DUE (downstream unwinding element).
П
 осле
синтеза РНК гетеродуплекс РНК-ДНК
оказывается прочнее, чем ДНК-ДНК, поэтому
РНК не вытесняется. На поздних стадиях
репликации вируса специфичность
РНК-полимеразы меняется.
осле
синтеза РНК гетеродуплекс РНК-ДНК
оказывается прочнее, чем ДНК-ДНК, поэтому
РНК не вытесняется. На поздних стадиях
репликации вируса специфичность
РНК-полимеразы меняется.

















Вирус кодирует РНК-ДНК-хеликазу UvsW (чувствительность к УФ-облучению). Она может расплетать РНК, синтезированную ДНК-матрице, поэтому на поздних стадиях размножения вируса инициации с участием РНК-полимеразы не происходит.
2.Рекомбинационная инициация
Рифампицин-резистентна.

















UvsX (аналог RecA E.coli) способствует внедрению одноцепочечной ДНК в двуцепочечный дуплекс. UvsY способствует посадке UvsX на ДНК. В процессе участвует также gp32 (аналог SSB E.coli). Этап миграции цепи осуществляется в определенном направлении 5'-3' по внедрившейся цепи, с затратой энергии АТФ. АТФ-азную реакцию катализирует UvsY. Возможно, АТФ нужен для перемещения. Размер "пузырька" постоянен, поэтому топологических затруднений для продвижения комплекса нет и топоизомераза не участвует.
Образование агрессивного 3' конца может происходить под действием эндонуклеаз. Внедриться может даже кольцевая молекула ДНК, но необходимо, чтобы был свободный 3' конец.
Свободный 3' конец используется полимеразой T4 в качестве затравки. Синтез ДНК осуществляется по консервативному механизму, как при синтезе РНК! Дочерняя цепь ДНК вытесняется из дуплекса. Размер репликативного глазка снова постоянен.
В синтезе отстающей цепи участвует gp61 – праймаза в комплексе с gp41 – хеликазой (5'-3'). Хеликаза олигомеризуется в кольцо. Для ее посадки нужен loader – gp59.




















Лидирующая цепь объединяется с остающей за пределами репликативного глазка. Переход от консерватизма к полуконсерватизму.
Специфичность праймазы:
pCpGppp
5'- dGdCdT -3'
pCpAppp
5'- dGdTdT -3'
Синтез РНК обычно начинается с пурина и обычно А.
Другие белки фага.
Топоизомераза фага состоит из трех субъединиц: gp39, gp52, gp60.
Экзонуклеаза (5'-3'): gp46, gp47.
Источники одноцепочечной ДНК.
1) недореплицированный 3' конец линейной ДНК
3' 5' 5'

3'

Из-за кольцевых пермутаций 3'конец одной молекулы может иметь гомологию с внутренним участком другой.
2)действие фаговой экзонуклеазы на случайных nicks.












Элонгация репликации.
Осуществляет ДНК полимераза фага. Это белок, называемый gp43 массой 103 кДа, состоящий из нескольких доменов. Фермент обладает также 3'-5' (proofreading) и 5'-3' (удаление РНК-затравки) экзонуклеазными активностями. Полимераза частично гомологична таковой у герпес-вируса. Сама по себе низкопроцессивна. Высокая процессивность полимеразы хороша при синтезе лидирующей цепи, но это становиться проблемой при прерывистом синтезе отстающей цепи. Фактором процессивности является гомотример белка gp45 (вспомните, гомодимер β-субъединицы полимеразы E.coli, гетеротример фактора PCNA эукариот). Аппарат загрузки фактора процессивности представлен комплексом одного белка gp44 и 4 копий gp42 (аналоги: γ-комплекс у прокариот и RF-C у эукариот).
Считается, что межсубъединичные связи в факторе процессивности неравноценны: одна слабая, поэтому при загрузке фактора кольцо можно разъединить по этой связи (с затратой АТФ).
Поскольку полимераза, праймаза и хеликаза образуют единый комплекс, то для объяснения одновременного синтеза лидирующей и отстающей цепей используют модель "трубы". То, что это единый комплекс узнали в опыте по разведению реакционной смеси, разведение не влияет ни на скорость синтеза ни на длину фрагментов ДНК.
Опыт по синтезу ДНК на матрице не содержащей С (???).
Удаление РНК затравок.
Фаговая РНКазаН удаляет РНК, фаговая полимераза застраивает брешь. У фага имеется Dda хеликаза (5'-3').
Препятствия на пути реплисомы создает встречная или догоняющая транскрипция. Происшествий не присходит: снимают шляпы и едут дальше. Другая проблема – прочно сидящий на ДНК белок.
Разрешение конкатемеров.
Вирусспецифическая резольваза (продукт гена VII) превращает ветвистые конкатемеры в неветвистые. Ее субстратом являются структуры Холлидея. Нарезание конкатемеров на мономеры сопряжено с упаковкой ДНК в вирусные частицы. Процесс осуществляет вирусная терминаза (гетеродимер gp16/gp17). Для упаковки нужна энергия АТФ. Происходит взаимодействие с одним из белков капсида – портальным белком.
Упаковка типа fullhead.
В ДНК обнаруживаются модифицированные основания: 5-оксиметилцитозин и гликозилированные производные нуклеотидов.
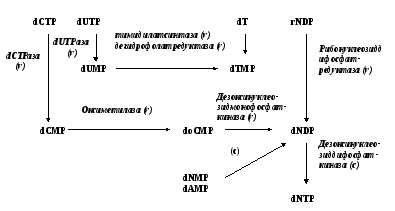
В экспериментах по включению субстратов синтеза нуклеиновых кислот в фаговую ДНК было установлено, что более ранние субстраты включаются в ДНК эффективнее, чем более поздние. При разрушении комплекса ферментов синтез ДНК останавливается.
Был сделан вывод о том, что ферменты образуют единый комплекс, в котором синтез нуклеотидов осуществляется по конвейерному принципу, в итоге нуклеозидтрифосфаты подаются прямо к активному центру полимеразы. Это позволяет использовать малые количества нуклеотидов.
OBP – центр кристаллизации всего комплекса. У EBV и герпесвирусов отсутствует OBP.