

Переход механизма репликации от схемы Кернса в схему разматывающего рулона.

Возможны несколько путей перехода.
1.Односторонняя репликация.




Гипотеза: превращение θ – σ, связано с ослаблением траскрипции в участке ori и, как следствие, возникновением односторонней репликации.
Причина ослабления транскрипции: а)действие cro-репрессора;
б) в клетке присутствует ограниченное количество белка DnaA, который влияет на транскрипцию с правого промотора фага. При увеличении количества фаговой ДНК, он вытитровывается и транскрипция ослабляется.
Если бы это был единственный механизм, то конец был бы фиксированным (как у φХ174). Но концы молекул могут разными.
2.Рекомбинационный механизм.
Фаг обладает red α,β,γ – системой: redα – 5’-3’экзонуклеаза, redβ – аналог SSB, redγ – ингибитор RecBCD системы (содержит экзонуклеазу), разрушающей линейные молекулы, а также σ-молекулы.
Redα образует агрессивный одноцепочечный 3’-конец, который внедряется в дуплекс кольцевой молекулы и вытесняет небольшой участок.
5’ 3’
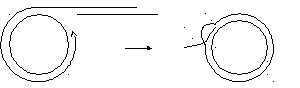
3.Внесение разрыва.
В принципе возможен случайный одноцепочечный разрыв.
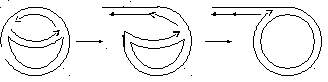
Механизмы не исключают друг друга и в принципе возможны.
Rep-хеликаза не участвует, поскольку белок О обладает хеликазной активностью.
В итоге образуются конкатемерные молекулы, которые надо разрезать на мономеры так, чтобы образовались липкие концы длиной 12 нуклеотидов. Нарезание осуществляется вирусной терминазой и сопряжено с упаковкой ДНК в вирусные частицы. Терминаза состоит из трех субъединиц А(Nu1)2 (А, потому что крайний слева в геноме). Разрезание происходит по специфическим участкам – cos.
Структура cos.








 CosQ CosN CosB
CosQ CosN CosB
I2 R3 I1 R2 R1

I1 , I2 - узнаются IHF (изгибает ДНК на 270˚С)
R1 , R2 , R3 - узнаются Nu1
Терминаза в виде димера присоединяется к CosB и на расстоянии 12 п.н. вносит разрыв в CosN. С этого участка начинается упаковка ДНК. CosQ нужен для правильного процессирования противоположного конца ДНК, чтобы упаковался точно один эквивалент генома. Поскольку для начала упаковки ДНК и завершения этого процесса используются сигналы одного и того же элемента, то должна быть регуляция специфичности внесения разрыва терминазой. Возможно, процесс регулируется путем взаимодействия белков капсида с терминазой.
Фаг р22 (лямбдоидный фаг).
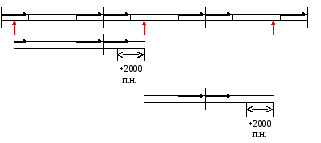
Геном представлен линейной двуцепочечной ДНК с тупыми концами. На концах прямые повторы длиной более 2000 п.н. Последовательность концевых повторов не фиксирована из-за кольцевых пермутаций. Причем пермутации полные, а у фага λ разброс в пределах 20%.

Как и у фага λ ori состоит из 4 итеронов, с прлегающей к ним А/Т-богатой областью. Ori находится внутри гена 18, гомологичного гену О, который кодирует аналог белка О фага λ. Рядом находится ген 12, кодирующий хеликазу.
На поздней стадии репликации трудно обнаружить кольцевые молекулы, но конкатемеры образуются. Может быть, конкатемеры образуются не по схеме Кернса.
Имеется белок угнетающий активность клеточной системы RecBCD. Упаковка ДНК начинается с сайта Pac разрыв вносится собственной терминазой. Работой упаковочной системы можно объяснить кольцевые пермутации. Имеется несколько Pac сайтов, разбросанных по геному. Pac сайт первого эквивалента генома выбирается случайно. Второй разрыв вносится после полного заполнения головки (fullhead упаковка). Помещается больше, чем эквивалент генома на длину прямого повтора (2000 п.н.). Упаковка следующего эквивалента генома начинается с конца первого (упаковка процессивна).
Фаг N15.
Открыт в России Виктором Равиным. Умеренный фаг. Геном представлен линейной двуцепочечной ДНК с липкими концами длиной 12 нуклеотидов. Левая половина генома как левая половина генома фага λ (структурные белки). В геноме закодирован белок RepA, функционально аналогичный белку A фага Р4, т.е. способен связываться с ori, обладает хеликазной и праймазной активностями.
В клетке происходит циклизация и сверхспирализация ДНК. Способен к лизогении, но есть потенциал и для репродуктивной инфекции.
Репродуктивная репликация изучена слабо, может быть, она осуществляется по схеме Кернса. Сейчас нас интересует репликация фага в лизогенном состоянии.
В состоянии профага существует в виде линейной плазмиды и не встраивается в геном клетки хозяина.
Внутри
генома есть сайт TelLR,
узнаваемый ферментом протеломеразой.
Протеломераза вносит двуцепочечный
разрыв в этот сайт с последующим
замыканием концов цепей ДНК. Образуется
структура похожая на ту, которую имеют
парвавирусы и поксвирусы, но в отличие
от них у фага N15 концы
комплементарны.
Инициация репликации. Репликация инициируется на участке ori белком RepA. Репликация двусторонняя, из-за ассимметричного расположения ori до одного из концов репликативная вилка дойдет раньше. Далее возможны два пути.
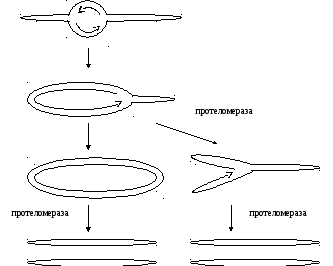
Механизм отнюдь не уникален. Недавно он был обнаружен еще у одного фага Ersinii и может осуществляться в клетке. Существует возбудитель Bardelia bardoferi, вызывающая болезнь Лайма, которая передается клещами. Ее хромосома имеет точно такую же структуру и реплицируется таким же механизмом.