

Что такое раздражимость, проводимость, возбудимость?
Всем без исключения живым тканям присуща раздражимость. Раздражимость – это универсальное свойство живых клеток отвечать на действие раздражителей изменениями структуры и функции, которые имеют неспецифический характер, а именно: изменением обмена веществ, теплообразования, роста и размножения клетки.
Возбудимость – это свойство высокоорганизованных (возбудимых) тканей реагировать на действие раздражителя изменением своего электрического состояния и специфическими функциональными проявлениями. В ответ на раздражение в возбудимых тканях возникает процесс возбуждения.
Возбуждение – это процесс генерации потенциала действия, его распространение по возбудимой ткани, приводящее к специализированному ответу. Оно характеризуется также изменением физических, химических, функциональных и структурных параметров клетки, но главное место занимает изменение электрического состояния: генерация потенциала действия, вслед за которым возникает деятельность, присущая данной ткани: мышца сокращается, нервная клетка генерирует нервный импульс, железа выделяет секрет. Сокращение мышц, генерирование нервных импульсов, выделение секрета – это специфические явления в тканях, сопровождающие процесс возбуждения. Кроме них всегда имеются и неспецифические проявления возбуждения – они такие же, как и во всех других тканях, не обладающих возбудимостью, а именно – это упомянутые ранее изменения обмена веществ, теплообразования, роста и деления клеток.
Возбудимость различных тканей неодинакова. Наиболее высокой возбудимостью отличаются рецепторы, затем следует нервная, мышечная и железистая ткани.
Проводимость – это свойство возбудимых тканей проводить волну возбуждения с определённой для данной ткани скоростью. В основе скорости проведения возбуждения лежит скорость биохимических реакций, протекающих на мембранах клеток.
Лабильность ткани (labilis, лат. – неустойчивый, скользящий). Это понятие ввёл в физиологию Н.Е.Введенский в 1901 г. для обозначения функциональной подвижности ткани. Под лабильностью понимают способность ткани отвечать на определённое ритмическое раздражение.
Мерой лабильности является максимальное количество импульсов, которое ткань способна воспроизвести в единицу времени без трансформации навязанного ритма.
Сократимость – это способность изменять механическое состояние сократительного аппарата цитоплазмы под влиянием раздражения.
Что такое раздражитель?
Раздражитель – это фактор внешней (внутренней) среды, который действует достаточно сильно, долго и нарастает с достаточной скоростью.
Раздражители классифицируют по нескольким признакам:
1) по модальности, т.е. по характеру энергии, свойственной раздражителю, они подразделяются на химические (кислоты, щелочи), осмотические, тепловые, электрические, световые, звуковые, словесные, биологические (медиаторы, гормоны, микробы);
2) по адекватности, т.е. по соответствию раздражителя воспринимающим раздражение рецепторам, они бывают адекватными и неадекватными. Например, адекватным раздражителем для рецепторов сетчатки глаз является свет, для барорецепторов – изменение давления и т.п. Пороговая сила адекватного раздражителя значительно ниже, чем неадекватного. Неадекватный раздражитель – это такой раздражитель, который действует на рецептор, не приспособленный для его восприятия. Например, возбуждение скелетной мышцы под воздействием не нервного импульса, а механического удара.
В физиологических опытах чаще всего используются электрические раздражители, т.к. они физиологичны, т.е. не повреждают ткань и напоминают нервные импульсы, и легко дозируются по силе, времени, крутизне нарастания (т.е. скорости нарастания силы тока во времени). Поэтому электрический ток принято рассматривать как адекватный раздражитель. Электрические раздражители по силе бывают:
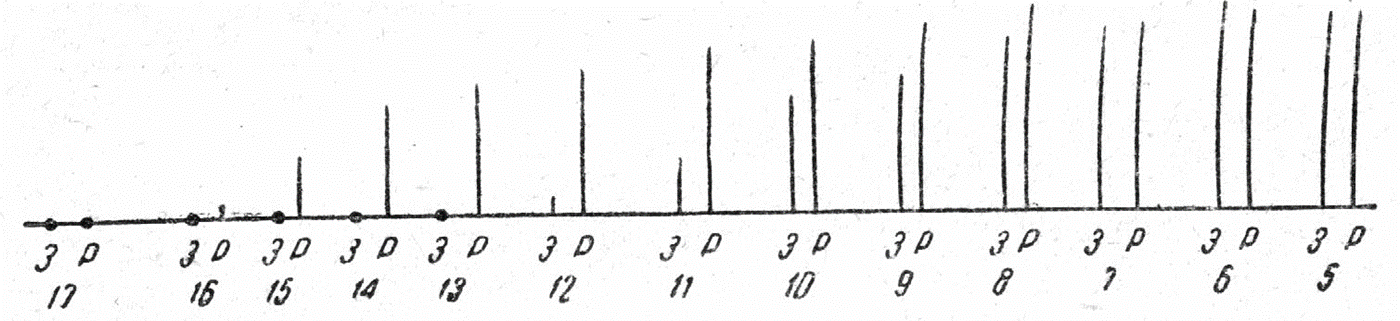
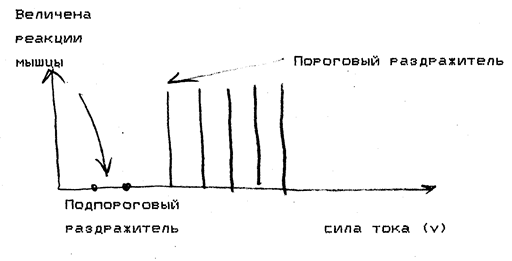
1) подпороговые раздражители – это слабые раздражители, не вызывающие видимой ответной реакции. В ткани возникает электротонический потенциал, а затем к нему присоединяется локальный ответ (местное возбуждение), что зависит от величины подпорогового раздражителя. Первые признаки локального ответа появляются при действии стимула, составляющего 50-75% от пороговой величины. Раздражители меньшей силы вызывают только электротонический потенциал;
2) пороговые раздражители – это минимальной силы раздражители, вызывающие генерацию потенциала действия и минимальный специфический физиологический эффект;
3) надпороговые раздражители – это более сильные раздражители, вызывающие пропорциональное увеличение физиологического эффекта;
4) максимальные раздражители – это раздражители такой силы, при которой физиологический эффект максимален;
5) супермаксимальные раздражители – это раздражители, величина которых больше максимальной, но эффект при этом не увеличивается. В процессе эволюции в трёх тканях – нервной, мышечной и железистой – раздражимость трансформировалась в новую, более совершенную форму реагирования ткани на действие раздражителей, а именно в возбудимость. Поэтому нервную, мышечную и железистую ткани называют возбудимыми.
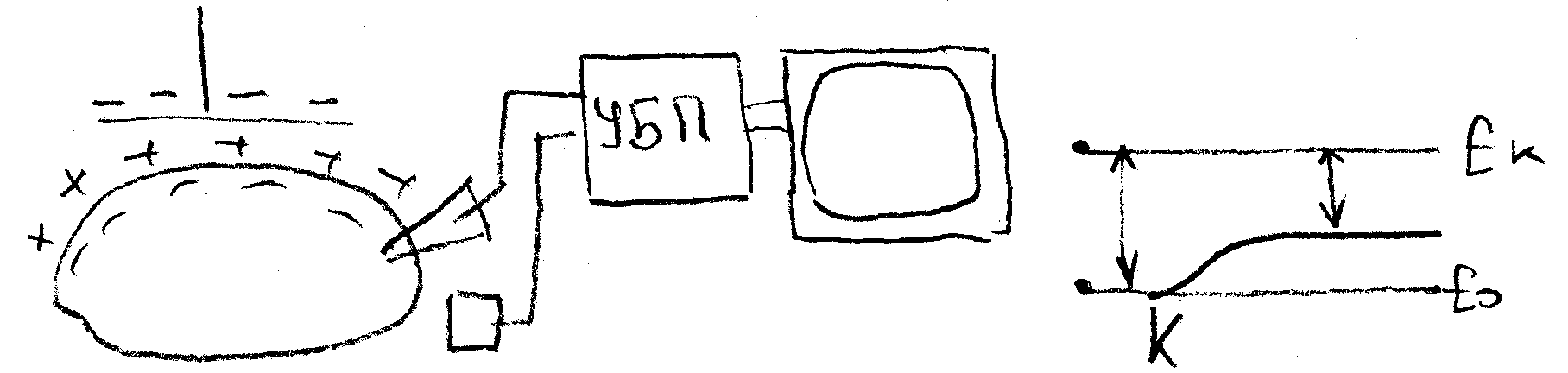
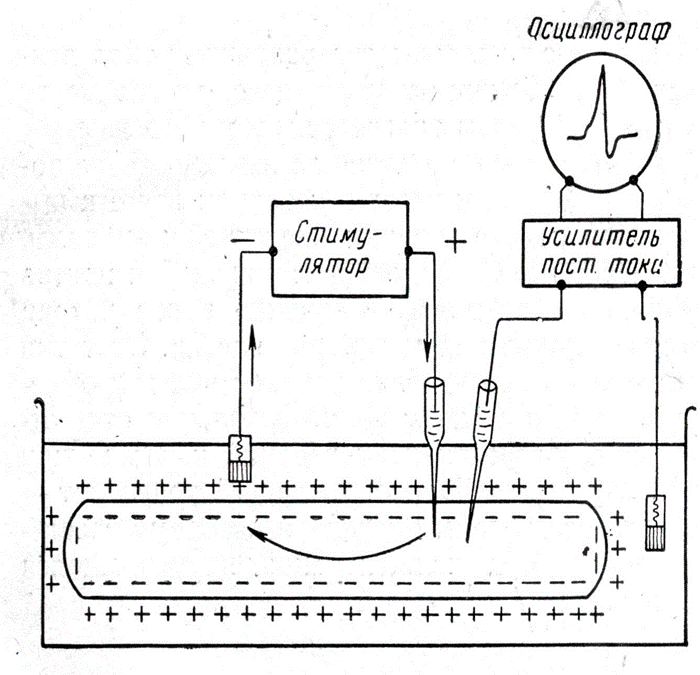
3. какое оборудование необходимо, чтобы зарегистрировать потенциал на мембране клетки?
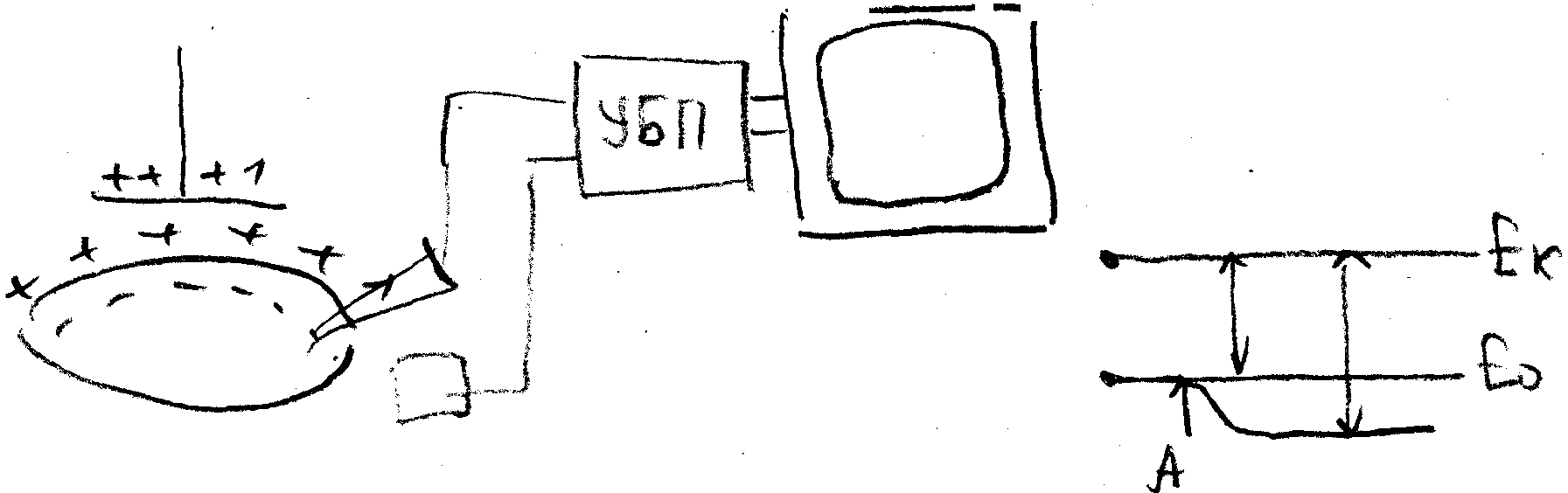
У всех живых клеток в покое плазматическая мембрана электрически поляризована, т.е. имеет разный электрический потенциал наружной и внутренней поверхностей. Это легко доказывается введением микроэлектрода, соединенного с усилителем и регистратором (осциллографом) внутрь клетки. Как только микроэлектрод проникает внутрь клетки, на экране осциллографа наблюдается скачок потенциала – от 0 до -70-80 мВ (по отношению к наружному электроду, расположенному в окружающей клетку жидкости). Эту величину называют мембранным потенциалом покоя или просто потенциалом покоя (ПП).
Прямое: внутриклеточная регистрация при помощи микроэлектрода
Непрямое (спектроскопическое): введение в клетку органических красителей и измерение их оптического сигнала (спектра флуоресценции или поглощения)
4.Опишите условия формирования мп и процессы, обеспечивающие его возникновение , в свете современной теории электрогенеза.
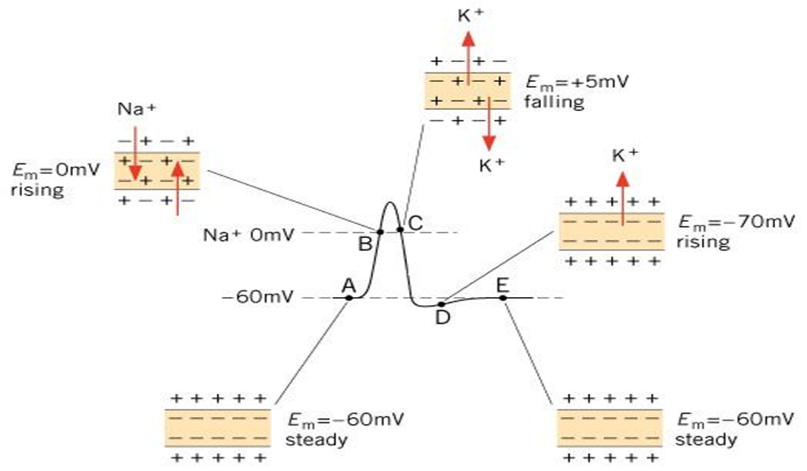
По концентрационному градиенту ионы калия выходят из клетки и заряжают наружную поверхность мембраны положительно. Крупные органические анионы заряжают внутреннюю поверхность мембраны отрицательно. На участке мембраны в 1 мкм 6 пар «+» и «-« ионов. Выход калия продолжается до установления равновесия – положительный заряд наружной поверхности мембраны и отрицательный заряд внутренней ее поверхности начинают препятствовать дальнейшему выходу положительно заряженного калия. Т.е. равновесный заряд – это заряд, препятствующий движению иона по концентрационному градиенту.
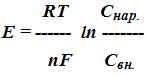
где Е - потенциал;
R - универсальная газовая постоянная, т.е кинетическая энергия 1 моля ионов при абсолютной температуре, равной 1о по Кельвину;
Т - абсолютная температура; n - валентность иона;
F - число Фарадея (заряд 1 моля одновалентных ионов);
Cнар. - концентрация ионов снаружи мембраны;
Свн. - концентрация ионов внутри клетки.
Установлено, что равновесие для ионов К+ в мышечном волокне теплокровных животных устанавливается при соотношении:
K,при этом Е К+ = -95 мВ;
для ионов Cl- при:
Cl, при этом Е Cl- = -90 мВ.
МП складывается из концентрационного потенциала и прямого электрогенного эффекта насоса.
Измерения в опыте потенциала покоя поперечно-полосатого мышечного волокна показало, что он равен -90 мВ, т.е. близок к Е К+.
В аксоне кальмара он равен -70 мВ (а по формуле Нернста Е К+ = -75 мВ, т.е. тоже близок к измеренному в опыте).
Если рассчитать по формуле Нернста потенциал в аксоне кальмара для Cl-, то он будет равен -90 мВ, т.е. он далёк от фактического потенциала, измеренного в опыте. Проницаемость мембраны для Cl- в нервных волокнах мала, и поэтому Cl- не играет существенной роли в генезе потенциала покоя. В скелетных мышечных волокнах проницаемость для Cl- сравнима с калиевой, и поэтому диффузия Cl- внутрь клетки увеличивает потенциал покоя. Аналогичная картина наблюдается для большинства клеток. Поэтому можно сделать заключение, что потенциал покоя обязан своим происхождением ионам К+, т.е. это калиевый равновесный потенциал.
Незначительное расхождение между величинами потенциала покоя, измеренными в опыте и рассчитанными по формуле Нернста, состоит в том, что в покое мембрана в небольшой степени проницаема для ионов Na+, и эти ионы, входящие внутрь клетки, уменьшают фактическое значение потенциала покоя. Поскольку мембрана тонкая потенциал покоя создаёт сильное электрическое поле напряженностью порядка 10 кВ/см.
Экспериментальное доказательство правильности такой точки зрения привели Ходжкин и Хаксли: они заменили аксоплазму в гигантском аксоне кальмара (диаметр 0,5-1,0 мм) на раствор КСl аналогичной концентрации, и в аксоне регистрировался потенциал покоя примерно такой же величины, как и в нативном нервном волокне.
Функцией мембранного потенциала покоя является действие электрического поля на макромолекулы мембраны, при этом заряженные группы этих молекул получают определенную пространственную ориентацию, и таким образом, например, обеспечивается закрытое состояние активационных ворот натриевых каналов и открытое состояние их инактивационных ворот. Этим самым обеспечивается состояние покоя и готовность к возбуждению. При дальнейшем местном возбуждении будет использоваться эта энергия, накопленная в потенциале покоя.
Теории электрогенеза
В 1896 году В.Ю. Чаговец предложил теорию электролитической диссоциации Арениуса применить для объяснения мембранных потенциалов
В 1902 году Ю. Бернштейн предложил мембранно-ионную теорию.
Современная теория (1949-1952 годы) предложена Ходжкиным, Хаксли и Кацем.
Основными положениями теории являются следующие:
· мембранный потенциал покоя - это диффузионный равновесный электрохимический потенциал;
· в возникновении мембранного потенциала покоя кроме ионов калия принимают участие и другие ионы ( натрий, кальций, хлор);
· в возникновении мембранного потенциала покоя вносит вклад электрогенный натрий-калиевый насос;
· потенциал действия определяется изменением полупроницаемости мембран. При возбуждении она становится хорошо проницаемой для натрия и перезаряжается.
5. что такое порог раздражения? Какова взаимосвязь порога раздражения и возбудимости ткани?
Порог раздражения - Минимальная сила раздражителя вызывающая развитие потенциала действия (специфический ответ возбудимой клетки)
Возбудимость клетки обратно пропорциональна порогу раздражения
Е=1/S
Порог раздражения (в физиологии нервных и мышечных клеток), наименьшая сила раздражителя (обычно электрического тока), способная вызвать распространяющийся потенциал действия; мера возбудимости клетки. В определённых пределах П. р. находится в обратной зависимости от длительности (t)действия стимула и крутизны нарастания его силы. С увеличением t П. р. падает. Только при некотором критическом увеличении t ("полезное время") П. р. устанавливается на постоянном уровне — реобазе. Минимальное значение t при силе тока, равной 2 реобазам, получило название хронаксии. При данном t величина П. р. одиночной клетки (волокна) зависит как от её "пассивных" свойств (сопротивление и ёмкость мембраны, сопротивление протоплазмы на единицу длины волокна), так и от активных свойств мембраны [состояние системы ионных каналов, в частности их чувствительность к деполяризации и скорость, с которой они способны активироваться (открываться) в ответ на деполяризацию].
Чтобы возникло возбуждение, раздражитель должен быть достаточно сильным – пороговым или сверхпороговым.
Учитывая, что порог раздражения является мерой возбудимости, которая определяется порогом деполяризации (разница между потенциалом покоя и уровнем критической деполяризации), то этот закон также должен рассматривать зависимость амплитуды ответа возбудимой ткани от силы раздражителя (раздражитель по силе ниже, равный или выше пороговой величины).
Для одиночных образований (нейрон, аксон, нервное волокно) эта зависимость носит название
Закон "все или ничего"
Подпороговые раздражители не вызывают возбуждение ("ничего"). При пороговых и сверхпороговых воздействиях возникает максимальная ответная реакция ("все"), т.е. возбуждение возникает с максимальной амплитудой ПД
По этому закону также сокращаются сердечная мышца и одиночное мышечное волокно.
Закон не являются абсолютным, а носит относительный характер:
1. При действии раздражителей подпороговой силы видимая реакция отсутствует, но возникает местная реакция (локальный ответ)
2. При действии пороговых раздражителей растянутая мышечная ткань дает большую амплитуду сокращения, чем не растянутая
При регистрации суммарной активности целостного образования (скелетная мышца, состоящая из отдельных мышечных волокон, нервный ствол, состоящий из множества нервных волокон)
проявляется другая зависимость: Чем больше сила раздражителя, тем больше величина ответной реакции.
6. назовите критерии возбудимости ткани.
Критерии оценки возбудимости ткани. Явление аккомодации возбудимой ткани. Параметры возбудимости ткани: пороговая сила (реобаза), полезное время, хронаксия. Функциональная лабильность ткани, мера лабильности. Изменение возбудимости при электротоническом изменении мембраного потенциала.
Возбудимость - это показать, который отражает насколько легко можно вызвать возбуждение.
Критериями оценки возбудимости ткани являются:
1. Порог деполяризации - это минимальная величина, на которую нужно деполяризовать клетку, чтобы вызвать ПД
2. Критический уровень ПД - это значение мембранного потенциала, при котором возникает ПД
Аккомодация - это снижение возбудимости при уменьшении крутизны ( скорости нарастания) раздражителя. Причина аккомодации такая же,как и при длительной деполяризации: при медленной деполяризации успевает развиться натриевая инактивация и калиевая активация.
Параметры возбудимой ткани:
1. Порог силы - это наименьшая сила, которую нужно приложить, чтобы вызвать ПД в неограниченное время.
2. Полезное время - это наименьшее время действия раздражителя на ткань (пороговой силы),чтобы вызвать ее возбуждение.
3. Хронаксия - это наименьшее время, в течение которого должен действовать электрических ток в 2 реобазы.
Лабильность - это скорость проведения ткани одного цикла возбуждения.
Мера лабильности - это количество циклов возбуждения, которое может провести ткань в 1 с.


7. опишите теории строения клеточной мембраны.
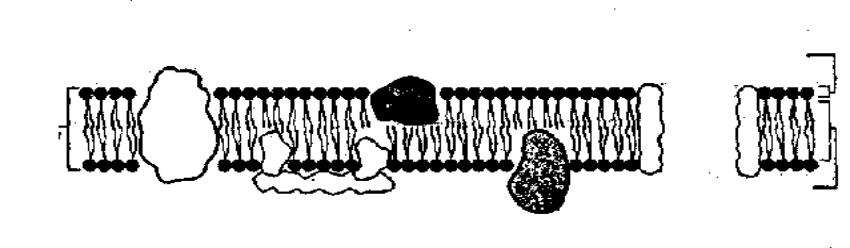
Ультраструктура биологических мембран.
Структурной основой биологической мембраны является двойной слой фосфолипидов, в который встроены мембранные белки.
Белки, пронизывающие насквозь фосфолипидный слой, называются внутренними мембранными белками, или белковыми каналами, или порами.
В функциональном отношении мембранные белки делятся на 4 класса: «насосы», каналы, рецепторы и ферменты.
«Насосы» расходуют метаболическую энергию АТФ для перемещения ионов и молекул против концентрационных и электрохимических градиентов и поддерживают необходимые концентрации этих молекул в клетке.
Ионоселективные каналы представляют собой пути переноса заряженных молекул и ионов. Через каналы в клетку проникают и лекарственные вещества.
Рецепторы мембран представлены белковыми молекулами, которые «узнают» то или иное биологически активное вещество, контактируют с ним и передают в клетку информацию о характере биохимических взаимодействий.
Белки-ферменты, обладающие высокой каталитической активностью, облегчают протекание биохимических реакций как внутри мембраны, так и у ее поверхности.
Современные представления о строении и свойствах клеточных мембран
Основу мембраны составляют липиды
МОЗАИЧНАЯ МОДЕЛЬ СТРОЕНИЯ МЕМБРАНЫ
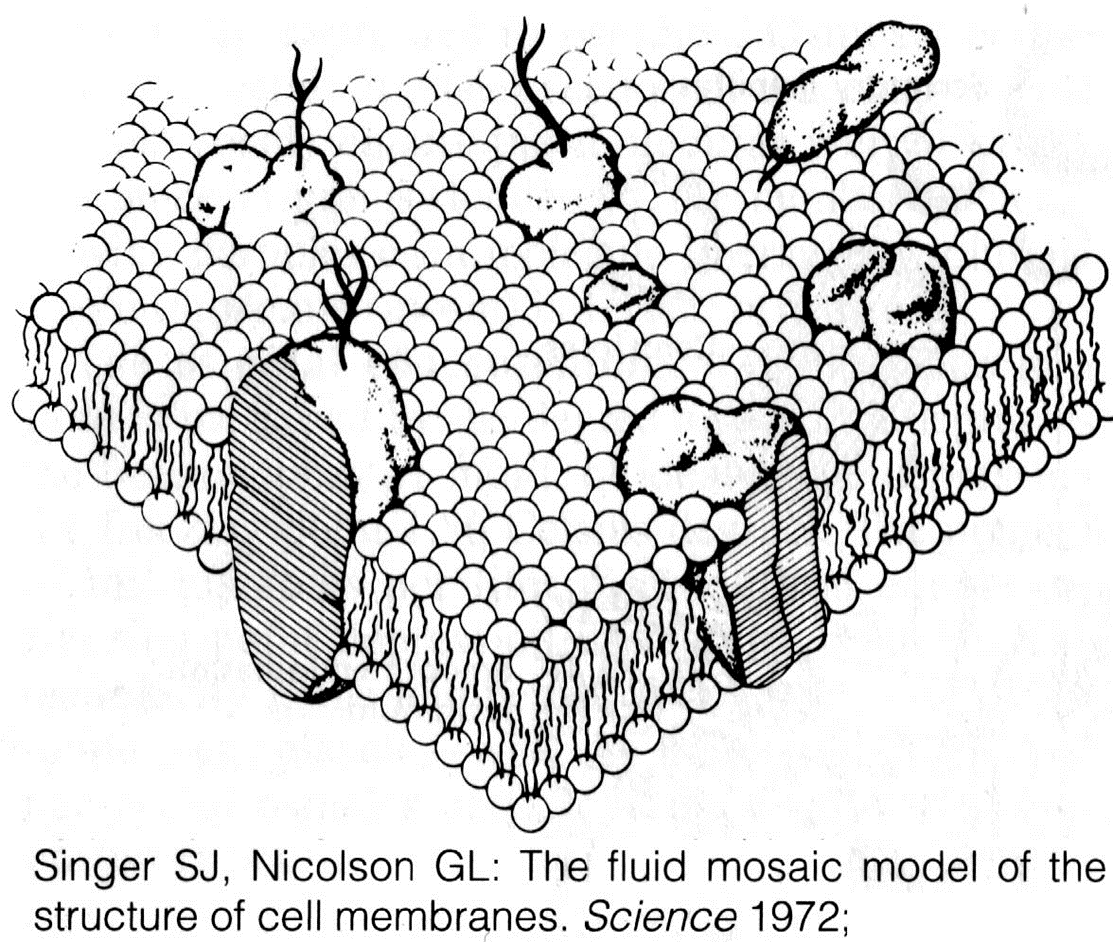
Современные представления о строении мембраны
Мембранные белки можно разделить на 2 типа.
Белки, прикрепленные к наружной поверхности мембраны с помощью электростатических сил. Их называют периферические белки.
Белки, пронизывающе билипидный слой, так, что одна часть глобулы погружена в мембрану, а другая в водную среду. Их называют интегральными. Некоторые из них свободно перемещаются в мембране, а другие крепятся к микротрубочкам и являются неподвижными.
С физиологической точки зрения белки всех клеток можно разделить на 5 классов:
каналы,
насосы,
рецепторы,
ферменты
и структурные белки.
Один и тот же белок может быть одновременно рецептором и каналом (ацетилхолин-стимулируемые каналы постсинаптической мембраны нервно-мышечного синапса) или ферментом и насосом (натрий-калиевая АТФ-аза).
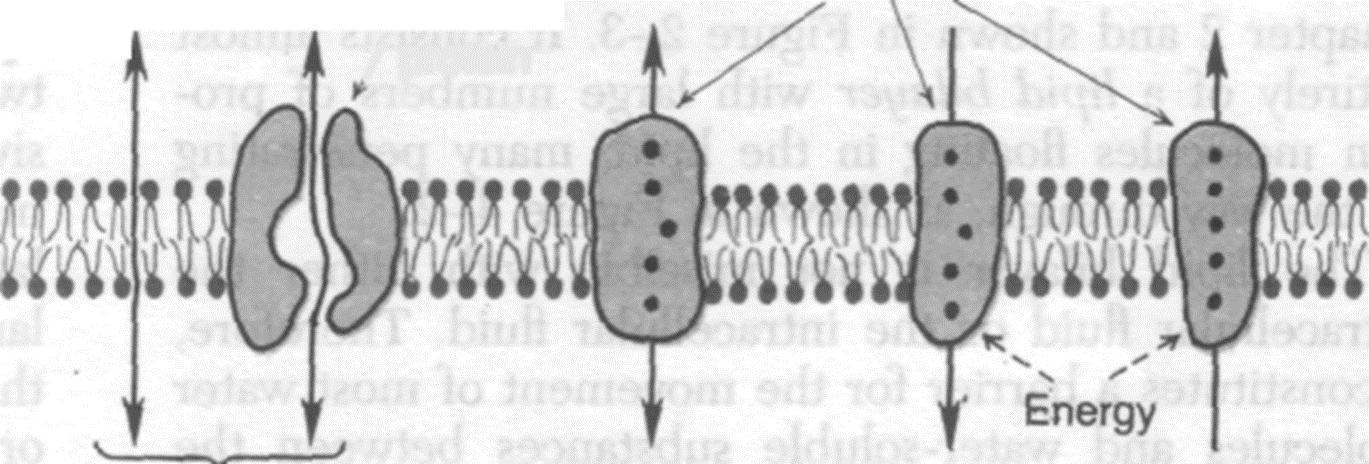
8. что такое пассивный и активный транспорт? Назовите виды пассивного траспорта веществ через мембраны клеток.
Основные виды ионного транспорта
Пассивный транспорт. Движение ионов осуществляется без использоваия энергии АТФ.
·Простая диффузия (движение ионов по градиенту концентрации).
·Осмос (движение растворителя по осмотическому градиенту).
·Электрохимический градиент (движение ионов в результате возникновения разности потенциалов).
Облегченная диффузия. (Возможна при участии белка переносчика)
Активный транспорт. Движение ионов осуществляется с использованием энергии АТФ против градиента их концентрации (Натрий-калиевый насос, кальциевый насос и различного рода переносчики).
Диффузия
Коэффициент диффузии — это мера скорости, с которой растворенное вещество может пересекать мембрану площадью 1 см2 и толщиной 1 см, когда разница концентраций вещества по разные стороны мембраны составляет 1 М/л.
Простая дифузия Облегченная Белки переносчики
Облегченная диффузия
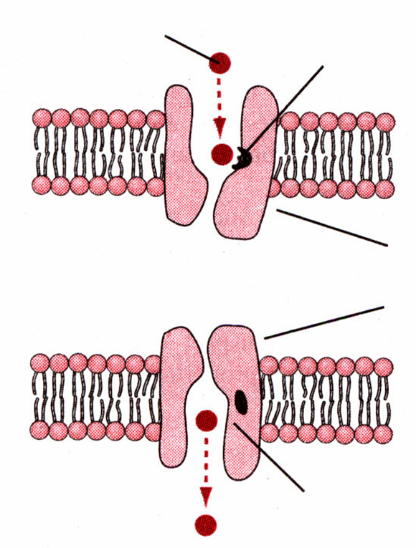 Связывающий
Связывающий
Сайт
Белок переносчик и
его конформационные
изменения
Отсоединение
От связывающего сайта
Пассивный транспорт. Фильтрация осуществляется через мембранные белковые каналы – поры, зависит от разности давлений снаружи и внутри клетки и проницаемости мембраны для жидкости и низкомолекулярных веществ. Диаметр пор чрезвычайно мал, поэтому фильтруются только низкомолекулярные вещества, вода и некоторые ионы.
Диффузия - пассивное передвижение молекул или ионов по градиенту концентрации (из области высокой концентрации в область низкой). Осмос представляет собой частный случай диффузии растворителя через полупроницаемую мембрану, не пропускающую растворенные вещества.
Пассивный транспорт не требует затрат энергии
9. что такое равновесный потенциал? Какова его величина для ионов К, Na, Cl?
Установлено, что равновесие для ионов К+ в мышечном волокне теплокровных животных устанавливается при соотношении:
K,при этом Е К+ = -95 мВ;
для ионов Cl- при:
Cl, при этом Е Cl- = -90 мВ.
МП складывается из концентрационного потенциала и прямого электрогенного эффекта насоса.
Измерения в опыте потенциала покоя поперечно-полосатого мышечного волокна показало, что он равен -90 мВ, т.е. близок к Е К+.
В аксоне кальмара он равен -70 мВ (а по формуле Нернста Е К+ = -75 мВ, т.е. тоже близок к измеренному в опыте).
Если рассчитать по формуле Нернста потенциал в аксоне кальмара для Cl-, то он будет равен -90 мВ, т.е. он далёк от фактического потенциала, измеренного в опыте. Проницаемость мембраны для Cl- в нервных волокнах мала, и поэтому Cl- не играет существенной роли в генезе потенциала покоя. В скелетных мышечных волокнах проницаемость для Cl- сравнима с калиевой, и поэтому диффузия Cl- внутрь клетки увеличивает потенциал покоя. Аналогичная картина наблюдается для большинства клеток. Поэтому можно сделать заключение, что потенциал покоя обязан своим происхождением ионам К+, т.е. это калиевый равновесный потенциал.
Незначительное расхождение между величинами потенциала покоя, измеренными в опыте и рассчитанными по формуле Нернста, состоит в том, что в покое мембрана в небольшой степени проницаема для ионов Na+, и эти ионы, входящие внутрь клетки, уменьшают фактическое значение потенциала покоя. Поскольку мембрана тонкая потенциал покоя создаёт сильное электрическое поле напряженностью порядка 10 кВ/см.
Экспериментальное доказательство правильности такой точки зрения привели Ходжкин и Хаксли: они заменили аксоплазму в гигантском аксоне кальмара (диаметр 0,5-1,0 мм) на раствор КСl аналогичной концентрации, и в аксоне регистрировался потенциал покоя примерно такой же величины, как и в нативном нервном волокне.
Функцией мембранного потенциала покоя является действие электрического поля на макромолекулы мембраны, при этом заряженные группы этих молекул получают определенную пространственную ориентацию, и таким образом, например, обеспечивается закрытое состояние активационных ворот натриевых каналов и открытое состояние их инактивационных ворот. Этим самым обеспечивается состояние покоя и готовность к возбуждению. При дальнейшем местном возбуждении будет использоваться эта энергия, накопленная в потенциале покоя.
Равновесный потенциал для иона — это такая величина мембранного потенциала, который установился бы по обе стороны клеточной мембраны, если бы она стала избирательно проницаема только для данного иона.

10. почему мембранный потенциал отличается от калиевого равновесного потенциала?
Установлено, что равновесие для ионов К+ в мышечном волокне теплокровных животных устанавливается при соотношении:
K,при этом Е К+ = -95 мВ;
11. напишите уравнение Нернста и укажите каково его значение.
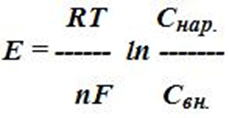
где Е - потенциал;
R - универсальная газовая постоянная, т.е кинетическая энергия 1 моля ионов при абсолютной температуре, равной 1о по Кельвину;
Т - абсолютная температура; n - валентность иона;
F - число Фарадея (заряд 1 моля одновалентных ионов);
Cнар. - концентрация ионов снаружи мембраны;
Свн. - концентрация ионов внутри клетки.
Уравнение Нернста — уравнение, связывающее окислительно-восстановительный потенциал системы с активностями веществ, входящих в электрохимическое уравнение, и стандартными электродными потенциалами окислительно-восстановительных пар.
Нернст изучал поведение электролитов при пропускании электрического тока и открыл закон. Закон устанавливает зависимость между электродвижущей силой ( разностью потенциалов ) и ионной концентрацией. Уравнение Нернста позволяет предсказать максимальный рабочий потенциал, который может быть получен в результате электрохимического взаимодействия, когда известны давление и температура. Таким образом, этот закон связывает термодинамику с электрохимической теорией в области решения проблем, касающихся сильно разбавленных растворов. E=E0+RTnFlnaOxaRed,
где
E — электродный потенциал, E0 — стандартный электродный потенциал, измеряется в вольтах;
R — универсальная газовая постоянная, равная 8.31 Дж/(моль·K);
T — абсолютная температура;
F — постоянная Фарадея, равная 96485,35 Кл·моль−1;
n — число молей электронов, участвующих в процессе;
aOx и aRed — активности соответственно окисленной и восстановленной форм вещества, участвующего в полуреакции.
Если в формулу Нернста подставить числовые значения констант R и F и перейти от натуральных логарифмов к десятичным, то при T=298K получим
E=E0+0,059nlgaOxaRed
12. опишите механизм возникновения потенциала действия.
Овершут
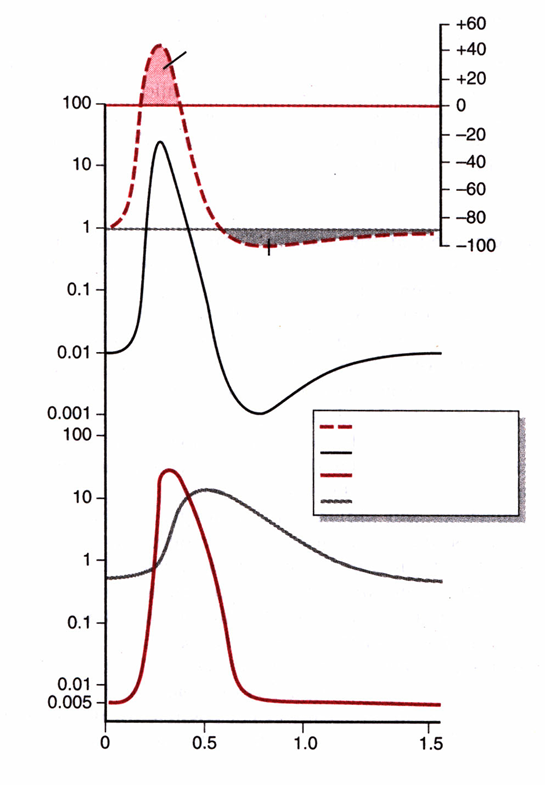
Следовая гиперполяризация Потенциал-зависимые натриевые каналы
Потенциал действия
Фазы потенциала действия
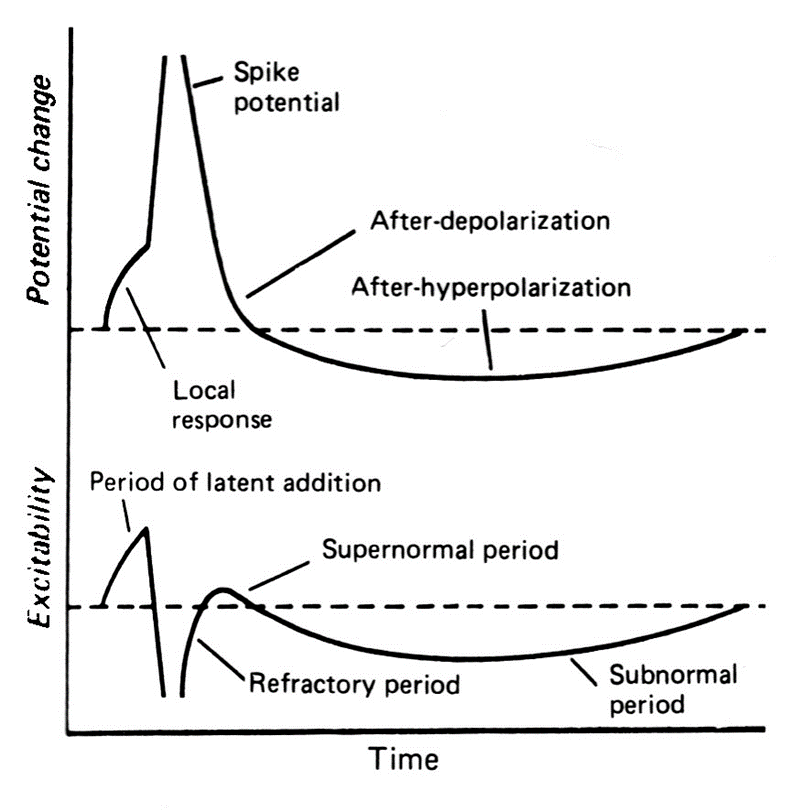
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
Отрицательный следовой потенциал — от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
Положительный следовой потенциал — увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
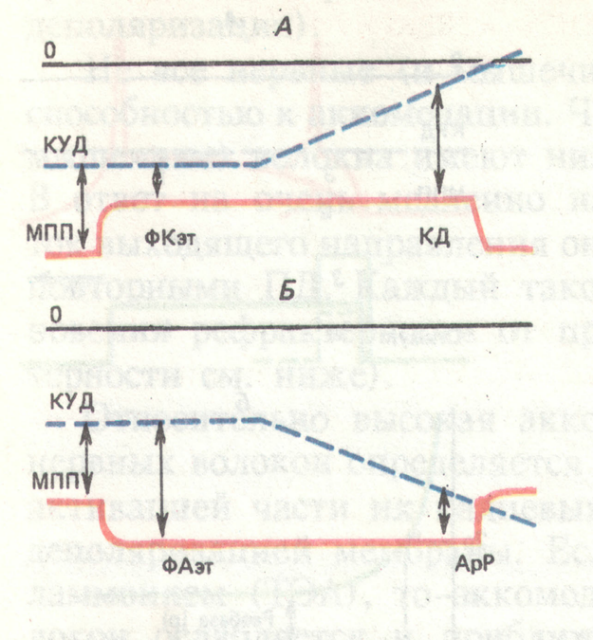
Фазы
ЛО- локальный ответ
Пик-пиковый потенциал
ПД-потенциал действия
СП-следовые потенциалы
КУД-критический уровень
деполяризации
МПП-мембранный
потенциал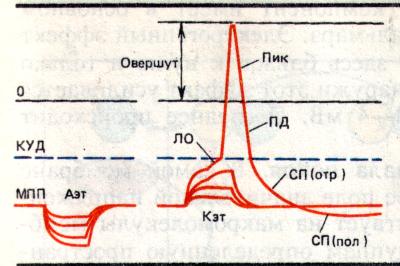
покоя
Аэт-анэлектротон
Кэт-катэлектротон
13. что такое деполяризация и каковы ее механизмы?
Уменьшение МП относительно его нормального уровня (ПП) называют деполяризацией
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциалчувствительных Na+-каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.

Рис. 2.4. Изменение мембран-ного потенциала, интенсивности калиевого и натриевого трансмембранного тока и возбудимости клетки в разные фазы потенциала действия.
Д – фаза деполяризации, Рб – фаза быстрой реполяризации, Рм – фаза медленной реполяризации, Г – фаза гиперполяризации;
Н – период нормальной возбудимости, Ра – период абсолютной рефрактерности, Ро – период относительной рефрактерности, Н+ – период супернормальной возбудимости, Н- – период субнормальной возбудимости
14. что такое порог деполяризации?
Порог деполяризации-это разность межу МП (Ео) и крити-ческим уровнем деполяризации (Ек):
Vt=Eo-Ek
Порог деполяризации является мерой возбудимости. Чем меньше порог деполяризации, тем меньшей силы необходимо при-ложить раздражитель чтобы вызвать развитие потенциала дейс-твия
15. в чем заключаются механизмы реполяризации?
фаза, во время которой восстанавливается исходный по тенциал покоя мембраны нервной клетки после прохождения через нее нервного импульса. Во время прохождения нервного импульса происходит временное изменение молекулярной структуры мембраны, в результате которого ионы могут свободно проходить через нее. Во время реполяризации ионы диффундируют в обратном направлении для восстановления прежнего электрического заряда мембраны, после чего нерв бывает готов к дальнейшей передаче через него импульсов.
Почему величина потенциала действия равна +40?
Это связано с тем, что та короткое время, в течение ко¬торого были от-крыты натриевые каналы и проницаемость мембра¬ны была увеличенной со-здается, равновесный диффузионный нат¬риевый потенциал.
Почему восстанавливается исходный мембранный потенциал после окончания потенциала действия?
Этот процессе состоит из 3 составляющих:
-инактивации натриевых каналов;
-увеличения проницаемости потенциал-зависимых калиевых кана-лов;
-работы натрий/калиевого насоса по восстановлению ион¬ных гради-ентов.
Инактивация натриевых каналов.
16. опишите механизм инактивации Na каналов.
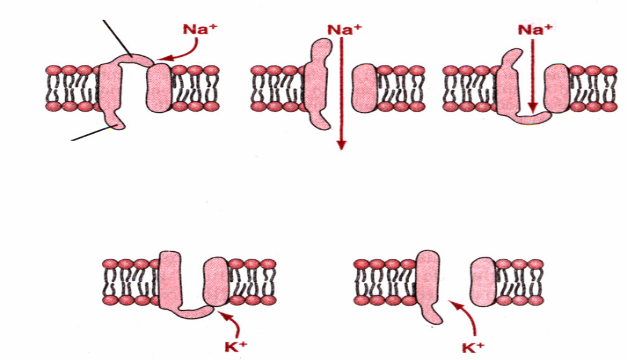
В потенциал-зависимых натриевых каналах имеется специ¬альное ворот-ное устройство. Одни из ворот располагаются бли¬же к наружной части канала и называются активационными М-воротами, а другие располагаются ближе к внутренней - инактивационные Н-ворота. В зависимости от состояния ворот-ных уст¬ройств можно выделить три состояния ворот: покоя, активации и инак-тивации.
В состоянии покоя активационные ворота закрыты, а инак-тивационные открыты. Через такие каналы ионы натрия прони¬кать не могут,
При деполяризации, когда мембранный потенциал уменьша¬ется от -90 до -70 или до -50, внезапно наблюдается конформационное изменение актива-ционных ворот и они открываются щелчком (то есть очень быстро). В это время проницаемость потенциал-зависимых натриевых каналов максимальна и проница¬емость мембраны для ионов натрия возрастает более чем в 500 раз. Следовательно, огромное количество ионов натрия посту¬пает в клетку. Это называется состоянием активации.
Далее наступает состояние инактивации натриевых кана¬лов. Изменение мембранного потенциала от -90 до -70- -50 мВ, которое привело к быстрому открытию активационных ворот, также является инициатором закрытия инак-тивационнных ворот. Однако закрытие инактивационных ворот происходит через 1/10000 секунды после открытия активационных ворот. Таким образом, конформационные изменения, сопровождающиеся откры¬тием активационных ворот - это процесс быстрый, а закрытие инактивационных ворот - медлен-ный, а открытие натриевых ка¬налов продолжается всего 1/10000 секунды.
17. назовите функции Na-K насоса.
Натрий-калиевый насос.
Натрий-калиевый насос вносит вклад в создание разности потенциалов на мембране клетки. Это возможно благодаря тому, что этот насос выводит ив клетки 3 положительно заряженных ионов натрия и вводит в клетку 2 по-ложительно заряженных иона калия. Это создает разность потенциалов на мембране приблизительно рав¬ной 4-10 мВ.
Таким образом, экспериментально полученный по¬тенциал покоя - 60 мВ и теоретически полученный - 60 мВ.
Значение натрий-калиевого насоса для жизни каждой клетки и организма в целом определяется тем, что непрерывное откачивание из клетки натрия и нагнетание в нее калия необходимо для осуществления многих жизненно важных процессов: осморегуляции и сохранения клеточного объема, поддержания разности потенциалов по обе стороны мембраны, поддержания электрической активности в нервных и мышечных клетках, для активного транспорта через мембраны других веществ (сахаров, аминокислот). Большие количества калия требуются также для белкового синтеза, гликолиза, фотосинтеза и других процессов. Примерно треть всей АТФ, расходуемой животной клеткой в состоянии покоя, затрачивается именно на поддержание работы натрий-калиевого насоса. Если каким-либо внешним воздействием подавить дыхание клетки, т. е. прекратить поступление кислорода и выработку АТФ, то ионный состав внутреннего содержимого клетки начнет постепенно меняться. В конце концов он придет в равновесие с ионным составом среды, окружающей клетку; в этом случае наступает смерть.
18. опишите зависимость силы ответной реакции от силы раздражителя ( закон силы ).
закон силы – возбудимость обратно-пропорциональна пороговой силе: чем больше пороговая сила, тем меньше возбудимость. Однако для возникновения возбуждения недостаточно только действия силы раздражения. Необходимо, чтобы это раздражение длилось какое-то время;
Для минимального возбуждения требуется определенная критическая пороговая сила действия раздражителя. Зависимость между силой раздражения и реакцией обьекта сформулировано в законе силы раздражения: чем сильнее раздражение, тем до известных пределов сильнее ответная реакция объекта (органа, ткани, клетки).
Под действием пороговых раздражителей суммарная активность целостного образования (мышцы или нервного ствола) градуально зависит от силы раздражителя в определен-ных пределах.
Скелетная мышца
19. в чем сущность закона «все или ничего». его критика.
Живая систе¬ма отвечает специфической реакцией на тот или иной раз-дражитель, если он обладает определенной силой. Если раздражитель имеет силу ниже этого уровня мышца на него специфической ре¬акцией не отвечает. Такие раздражители называются подпороговыми.
Дальнейшее увеличение силы раздражителя сопровождается уве-личением силы ответной реакции.
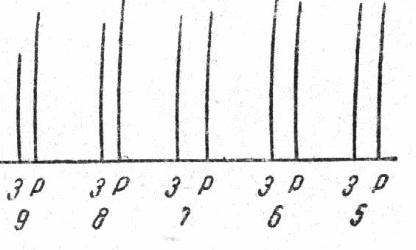
Та минимальная сила тока, на которую мышца отвечает макси-мальным сокращением называется максимальным раздражите¬лем. В 1871 г. Боудич изучал закон силы на сердечной мышце, и обнаружил, что мышца сердца на подпороговое раздражение не отвечает сокраще-нием, а на пороговое отвечает максимальным сокращением и при дальнейшем увеличении силы раздражителя сила сокращения мышцы не изменяется
Этот закон получил название "Все или ничего"
Таким образом , скелетная мышца отвечает градуальной реак-цией < чем больше, тем больше), а сердечная мышца подчи¬няется зако-ну " Все или ничего". Боудич распространил закон "Все или ничего" на все возбудимые ткани.
 сердечная
мышца
сердечная
мышца
Почему скелетная мышца отвечает градуальной реакцией, а сер-дечная мышца подчиняется закону "Все или ничего"?
Любая скелетная мышца состоит из большого количества мы-шечным волокон. Разные волокна не одинаково реагируют на раздра-жители, т.е. они обладают разной возбудимостью. По ме¬ре нарастания силы тока в процесс включаются все новые и но¬вые мышечные волокна, и сократительная реакция скелетной мышцы увеличивается. В сердеч-ной мышце каждая клетка имеет приблизительно равный порог раз-дражения, поэтому при нанесении порогового раздражителя все кар-диомиоциты сокращаются одновременно. Второй причиной является то, что сердечная мышца представля¬ет собой функциональный синци-тий.
КРИТИКА ЗАКОНА " ВСЕ ИЛИ НИЧЕГО "
Критика положения «ничего». Боудич (1840-1911) предполагал, что "ничего" означает полное отсутствие ответа на подпороговый стимул, а "все" рассматривал, как максимально возможную реакцию ткани. Ухтомский возражал Боудичу и говорил, что любая клетка подчиняется закону силы, а не закону "Все или ничего". Несмотря на то, что на подпороговые раздражители не возникает сокращение, разность потенциалов на мембране изменяется. Вот это изменение мембранного потенциала подчиняется закону силы.
Критика положения "все". Максимально возможный ответ находится в прямой зависимости от температуры, исходного растяжения и т.д. Закон "Все или ничего" относителен, а не абсолютен, как это полагал Боудич. Этот закон вторичен, т.е. неспецифические компоненты перерастают в специфические физиологические ответы.
Графически зависимость сила-время математически выража¬ется гиперболой.

a
I = ----- + b ;
t
где I – сила тока;
t – время действия тока;
a и b – постоянные величины.
-сила тока в толчках очень малой величины
t-время действия раздражителя
Ь-реобаза.
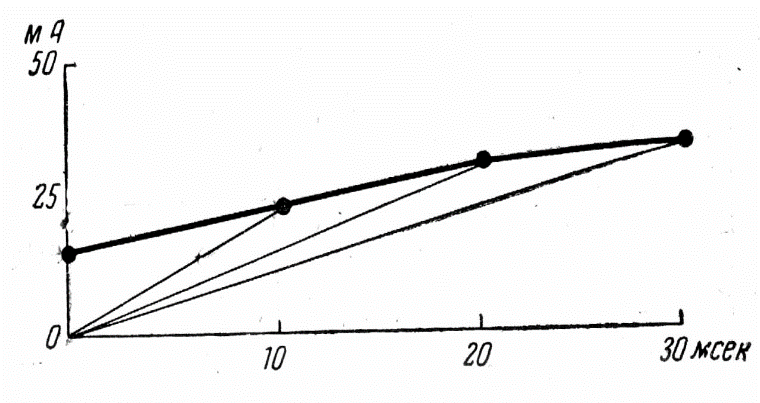
20. в чем сущность закона длительности раздражения? Нарисуйте кривую Гоорвега-Вейса.
Закон сила – время (кривая Горвега-Вейса)
Закон
устанавливает зависимость между силой
раздражителя и временем его действия:
«Пороговая сила любого раздражителя
обратно пропорцио-нальна длительности
его действия в определенных пределах».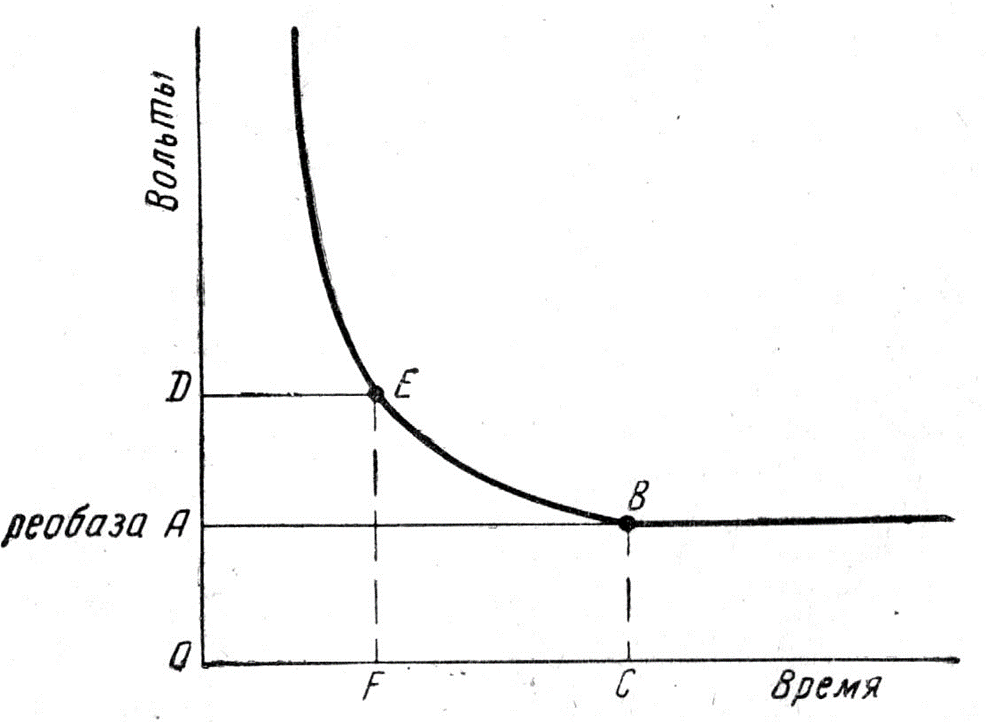
21. что такое полезное время и хронаксия?
Закон сила – время
Наименьшее время, которое должна действовать реобаза, чтобы вызвать возбуждение, называется полезным временем.
Хронаксия – это время, которое должен действовать ток, равный по величине удвоенной реобазе, чтобы вызвать возбуждение.
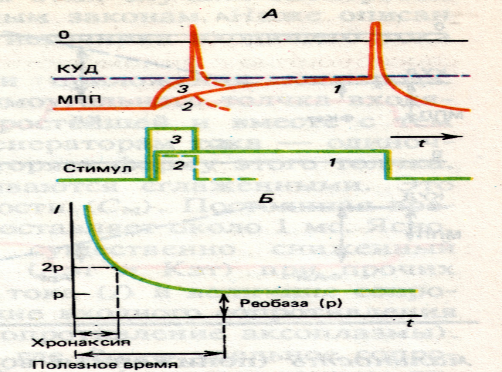
хроноксия- это наименьшее время, в течение которого электрический ток, равный по силе удвоенной реобазе, должен действовать на ткань, чтобы вызывать возбуждение. хроноксия бывает чувствительная и двигательная.
Для определения кроноксиии применяют специальные приборы-хроноксиметры.
22. что такое хронаксиметрия
При помощи этих приборов измеряют возбудимость мышц и нервных стволов. Путем измерения мроноксии невропатолог может определить повреждение нерва.
вторым местом приложения закона "Сила-время" являются аппараты УВЧ (ультравысокой частоты). Это генераторы, которые создают ток по величине несколько тысяч вольт” но длительность одного импульса чрезвычайно мала. В связи с этим ток оказывает только тепловое воздействие.
В неврологической практике проводят хронаксиметрию, чтобы установить поражение двигательных нервов. Если нерв повреждён или погибли мотонейроны спинного мозга (полиомиелит), то нервные волокна перерождаются и раздражающий стимул выявляет увеличение реобазы и хронаксии.
23. в чем сущность закона градиента раздражения?
Для того чтобы раздражитель вызвал генерацию ПД, его сила должна нара-cтать достаточно быстро. При какой-то минимальной крутизне нарастания интенсивности раздражения (минимальный градиент) ответы на это раздражение исчезают, т.к. в ткани развивается процесс аккомодации (accomodation, англ. – приспособление). Величина минимального градиента, выраженного в единицах реобаза в секунду (МА), является показателем скорости аккомодации.
24. что такое аккомодация ткани?
при уменьшении крутизны нарастания тока ниже некоторой минимальной величины потенциал действия вообще не возникает, до какой бы силы не увеличивали ток. Такое явление приспособления возбудимой ткани к медленно нарастающему раздражителю и называется аккомодацией.
Аккомодация обусловлена тем, что за время действия электрического тока в мембране успевают развиться процессы, препятствугощие возникновению потенциала действия.

Аккомодация выражается в единицах реобаза в секунду. С точки зрения электрофизиологии в основе аккомодации лежат активация натриевой и повышение калиевой проводимости ,развивающееся во время медленно нарастающей деполяризации мембраны.
25. в чем сущность и каковы основные положения полярного закона?
ПОЛЯРНЫЙ ЗАКОН РАЗДРАЖЕНИЯ.
при раздражении нерва или мышцы постоянным током посредством приложенных электродов, приложенных к поверхности нерва или мышцы!
1>возбуждение возникает в момент замыкания цепи и в момент размыкания цепи*
2) в момент замыкания возбуждение возникает под катодом, а в момент размыкания под анодом*
3)раздражающее действие катода сильнее, чем анода.
Эти факты объединяют под названием «Полярный закон». Этот закон был открыт Пфлюгером в 1859 г.
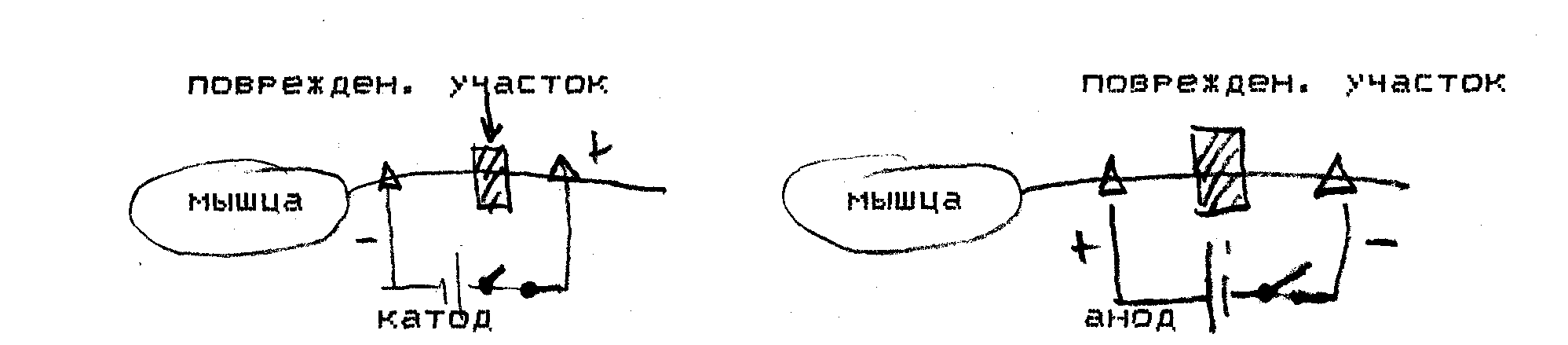
1. Возбуждение и сокращение 11.Возбуждение возникает в момент мышцы возникает в момент размыкания цепи замыкания эл. цепи.
Физический электротон ~ это изменение мембранного потенциала под влиянием постоянного тока.
Можно дать другое определение '•
Физический
электротон - это изменение мембранного
потенциала не связанное с изменением
проницаемости мембраны для ионов.
<——"j„
•••
',
В области, где приложен к ткани анод (положительный по- люс) ,, положительный потенциал на наружной поверхности мемб- раны возрастает, то есть происходит гиперполяризация.
В месте воздействия отрицательного полюса постоянного тока - катода - положительный потенциал на наружной поверхности мембраны уменьшается, то есть происходит деполяризация.
Изменение мембранного потенциала возникает не только непосредственно в точках приложения к нервному волокну катода и анода постоянного тока. Они наблюдаются также и на некотором расстоянии от полюсов с той, однако” разницей, что v\x величина постепенно убывает по мере удаления от катода и анода.
Вызываемые постоянным током изменения мембранного потенциала не связаны с изменением ионной проницаемости мембраны клетки, то есть с ее активным ответом на приложенный ток. Поэтому их принято называть пассивными сдвигами потенциала s отличие от активным его изменений - активной деполяризации и активной гиперполяризации мембраны, возниканяция при возбуждении и обусловленные! изменением проницаемости этой мембраны.
Католическая депрессия Вериго- при действии постоянного тока под катодом развивается снижение возбудимости, а под анодом возбудимость возрастает.
Физиологический электротон- это изменение возбудимости мембраны нервной или мышечной клетки под полюсами постоянного тока. Под катодом при замыкании происходит повышение возбудимости, а под анодом ее снижение (катэлектротон и анэлект-рои) .
26. что такое физический и физиологический анэлектротон?
физиологический (ан- + электротон) понижение возбудимости и проводимости возбудимой ткани в области анода при действии на ткань постоянного тока. Физический анэлектротон – повышение МП
27. что такое физический и физиологический катэлектротон?
Физиологический катэлектротон – повышение возбудимости за счёт снижения МП при «нормальном» КУД
Катодическая депрессия - снижение возбудимости при низком МП за счёт повышения уровня КУД
28. какие изменения возбудимости вызываются изменением мп?
Изменение мембранного потенциала возникает не только непосредственно в точках приложения к нервному волокну катода и анода постоянного тока. Они наблюдаются также и на некотором расстоянии от полюсов с той, однако” разницей, что v\x величина постепенно убывает по мере удаления от катода и анода.
Вызываемые постоянным током изменения мембранного потенциала не связаны с изменением ионной проницаемости мембраны клетки, то есть с ее активным ответом на приложенный ток. Поэтому их принято называть пассивными сдвигами потенциала s отличие от активным его изменений - активной деполяризации и активной гиперполяризации мембраны, возниканяция при возбуждении и обусловленные! изменением проницаемости этой мембраны.
29. какие изменения возбудимости вызываются сдвигом критического уровня деполяризации?
Критический уровень деполяризации
Если регистрировать изменение мембранного потенциала нервного волокна, то потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического уровня. Критический уровень деполяризации не зависит от ка-рактера раздражителя, а определяется исключительно свойствами мембраны.


-60
Vt=Eo-Ek

-70
Порог деполяризации – это разность между мембранным потенциалом покоя (Eo) и критическим уровнем деполяризации (Ek).
Порог деполяризации является мерой возбудимости. Чем меньше порог деполяризаци, тем меньшей силы необкодимо приложить раздражитель чтобы вызвать развитие потенциала действия.
30. перечислите фазы изменения возбудимости при возбуждении.
Возникновение в нервном или мышечном волокне потенциала действия сопровождается многофазными изменениями возбудимости.

Во время локального ответа возбудимость повышается.
Периоду развития пикового потенциала соответствует-полное исчезновение возбудимости (абсолютная реФрактерность). В это время второе раздражение не способно вызывать новый потенциал действия ( он длиться 0,4 с в нервном волокне).
Вслед за абсолютной реФрактерностью начинается Фаза относительной реФрактерности. В это время возбудимость постепенно восстанавливается.
Относительная реФрактерность сменяется супернормальным периодом, т.е. периодом повышенной возбудимости.
Затем развивается субнормальный период. Этому периоду соответствует снижение возбудимости.
31. что такое абсолютный рефрактерный период?
Интервал, в течении которого возбудимая ткань не способна генерировать потенциал действия, каким бы сильным ни был раздражитель.
32. что такое распространяющееся возбуждение и его характеристика?
При достижении местным возбуждением некоторой пороговой величины (порога раздражения) возникает потенциал действия (ПД) — распространяющееся возбуждение.
Пик потенциала представляет собой кратковременную ин¬версию внут-риклеточного потенциала. В естественных условиях ПД возникает локально, а затем распространяется, то есть ПД есть распоястраняющееся возбуждение
Возникает на пороговый или надпороговый раздражитель, не способно к суммации, подчиняется закону «все или ничего», т.е. если местное возбуждение не достигает критического уровня деполяризации, то ПД не возникает, а если достигает – то ПД возникает с одинаковой амплитудой, зависящей только от свойств возбудимой ткани. возбудимость снижается, продолжительность возб менее длительна, чем местное возбуждение. Распространяется от места возникновения по возбудимой ткани без декремента
33. что такое местное возбуждение и его особенности?
ПРИ нанесении слабого ПОДПОРОГОВОГО раздражителя разви¬вается сла-бый локальный ответ или местный потенциал. Локаль¬ным он называется по-тому, что и в экспериментальных и ес¬тественных условиях этот потенциал не распространяется дале¬ко.
Свойства местного возбуждения
• Возникает на подпороговый раздражитель. По мере даль¬нейшего уси-ления раздражающего тока локальный ответ увеличи¬вается и в момент , ко-гда деполяризация мембраны достигает критического уровня-возникает по-тенциал действия.
• Местные потенциалы суммируются.
• Местные потенциалы (возбуждение) не ПРИВОДЯТ к воз¬никновению спе-цеФической реакции.
• В момент возникновения местного возбуждения возбуди¬мость увеличи-вается.
• Местные потенциалы подчиняются закону силы, а не за¬кону "Все или ничего".
34. что собой представляют активные и пассивные сдвиги потенциала?
Сдвиги мп произошли пассивно – за счет энергии раздражителя
Изменение мп под катодом – пассивная катодическая деполяризация (катэлектротон)
Изменение мп под анодом – пассивная анодическая гиперполяризация (анэлектротон)
Это физический электротон - изменение мп, возникающее по влиянием постоянного тока, без изменения проницаемости мембраны для ионов.
Пассивные сдвиги мп (электротонические потенциалы) возникают за счет энергии раздражителя, не связаны с изменением проницаемости мембраны для ионов, развиваются постепенно( это зависит от емкости и сопротивления мембраны), находятся в градуальной зависимости от силы раздражителя
В естественных условиях причиной возникновения потенциала действия (ПД) является местный ток, который возникает между возбужденным и покоящимся участками мембраны.
В процессе возбуждения ткани в ней происходят сдвиги мембранного потенциала, вначале они носят пассивный характер, а затем активный.
Пассивные сдвиги потенциала возникают в ответ на действие любого раздражителя за счет его энергии. В отчетливой форме они лучше всего выявляются при действии полюсов постоянного тока (катода и анода).
Пассивные изменения мембранного потенциала развиваются от момента нанесения раздражения до возникновения местного возбуждения.
Физический электротон - это изменение мембранного потенциала, возникающее под влиянием постоянного тока и не связанное с изменением проницаемости мембраны для ионов.
Катэлектротон
Анэлектротон
Электротонические потенциалы:
нарастают и исчезают постепенно;
способны к суммации;
распространяются с декрементом;
при увеличении подпорогового тока до 50% от порогового значения возникает активный сдвиг потенциала в виде местного возбуждения.
Активные сдвиги – локальный ответ( местное возбуждение) и пд (распростр возбужд)
Гиперполяризующие толчки тока (длительностью 4 мс) с относительной амплитудой 1, 2, 3, 4 и 5 вызывают пропорциональные по амплитуде электротонические потенциалы. При деполяризующих токах с амплитудой 1 и 2 возникающие потенциалы являются зеркальным отражением потенциалов при соответствующих гиперполяризу-ющих токах. При деполяризующих толчках с амплитудой 3 и 4 электротонические потенциалы выше тех, которые возникают при деполяризации менее - 70 мВ (область превышения затушевана розовым). Ток с амплитудой 5 производит деполяризацию, которая превышает порог и вызывает потенциал действия.
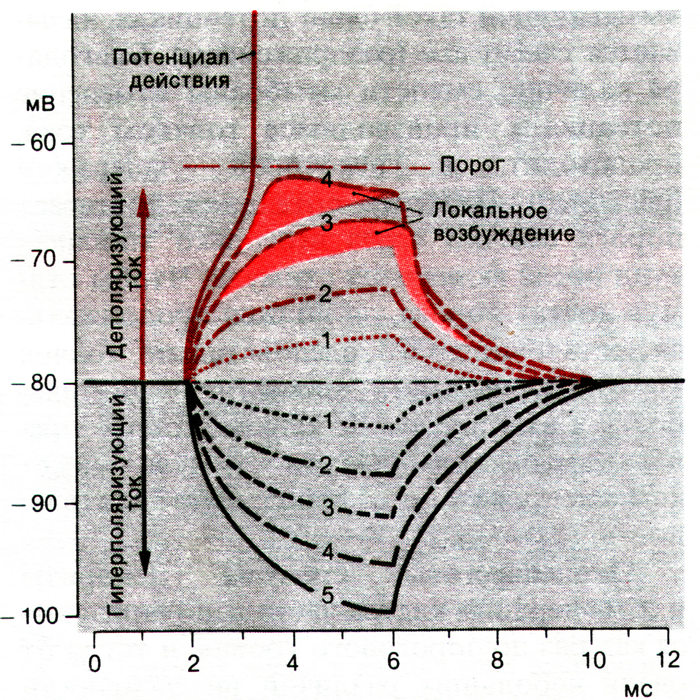
На кривых, близких к пороговому уровню, (на рис. затушевана розовым) деполяризация называется локальным ответом и обусловлена повышением Na+-проводимости. Во время локальных ответов вход Na + может существенно превосходить выход К +, однако Na + -ток еще не так велик, чтобы деполяризация мембраны стала достаточно быстрой для генерации потенциала действия или возбуждения соседних участков ткани. Возбуждение развивается не полностью, остается локальным процессом и не распространяется.
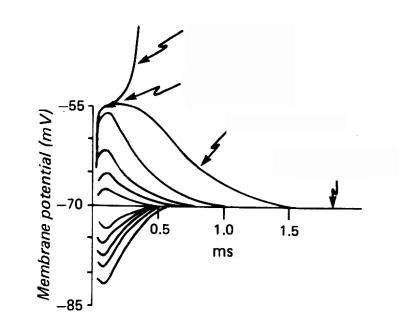
Если регистрировать изменение мембранного потенциала нервного волокна, то потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического уровня. Критический уровень деполяризации не зависит от характера раздражителя, а определяется исключительно свойствами мембраны.
Порог деполяризации является мерой возбудимости. Чем меньше порог деполяризации, тем меньшей силы раздражитель необходимо применить для того чтобы вызвать потенциал действия.
Vt=Eo-Ek,
Vt – порог деполяризации
Ео – величина потенциала покоя
Ек – величина критического уровня деполяризации
Физиологический электротон это изменение возбудимости мембраны нервной или мышечной клетки под полюсами постоянного тока. Под катодом при замыкании происходит повышение возбудимости, а под анодом ее снижение (катэлектротон и анэлектротон).
35. в чем заключается физиологическое значение мышц?
Обеспечивают перемещение тела в пространстве, взаимодействие организма с окружающей средой, работу внутренних органов.
36. что такое прямое и непрямое раздражение мышцы?
При непосредственном приложении раздражителя к мышце, или прямом раздражении, можно вызвать ее сокращение и после перерождения двигательного нерва, после отравления мионевральных аппаратов ядами, прекращающими передачу возбуждения с нервных волокон на мышечные. При непрямом раздражении, когда раздражитель действует на двигательный нерв, достаточен более слабый раздражитель, так как возбудимость нерва больше, чем мышцы.
37. как определяется работа и сила мышцы? От чего они зависят?
Закон средних нагрузок.
А = Р х Н, где
А - работа,
Р - груз, поднимаемый на высоту - Н.
Работа выражается в кгм и гсм.
Способность выполнять работу и поднимать тот или иной груз зависит от силы мышцы.
Сила мышцы - это величина максимального груза, который мышца еще может поднять при максимальном изометрическом напряжении. Сила мышцы зависит и от физиологического поперечного сечения мышцы.
Абсолютная сила мышцы – это частное от деления силы мышцы на 1 см 2 её площади.
Правило средних нагрузок
Между величиной нагрузки и производимой мышечной работой существует определенная зависимость: по мере увеличения нагрузки мышечная работа возрастает до определенного уровня, а затем уменьшается. Максимальная мышечная работа производится при средник нагрузках - правило средних нагрузок.
38. коэффициент полезного действия мышцы.
В ходе изотонического мышечного сокращения выделение тепла пропорционально выполненной работе. Коэффициент полезного действия мышечной работы (КПД) представляет собой отношение величины внешней механической работы (А) к общему количеству выделенной в виде тепла (Е) энергии.
Наиболее высокое значение КПД (R) мышцы наблюдается при внешней нагрузке, составляющей около 50% от максимальной величины.
Величена КПД в машинах 15%, в живой мышце - около 25-35%. Отличие мышцы от неживой машины:
1) в мышце используется для работы не тепловая, а хими¬еская энергия
2) в процессе работы мышца не изнашивается, а наоборот, совершенствуется и сила мышцы возрастает.
39. назовите виды сокращения мышц?
Одиночное мышечное сокращение. Раздражение мышцы или иннервирующего ее двигательного нерва одиночным стимулом вызывает одиночное сокращение мышцы. Одиночное сокращение имеет вид волны.
Тетаническое мышечное сокращение. Мышца может быть возбуждена еще во время сокращения - при ритмическом раздражении.
Тетанус - слитное сокращение, возникающее при ритмической стимуляции, когда время между сокращениями меньше, чем длительность одиночного сокращения. В связи с тем, что во время сокращения мышца не имеет рефрактерности, повторные стимулы, наносимые до расслабления, способны создавать дополнительную активацию сократительных элементов и увеличивать силу мышечного сокращения. Этот феномен носит название гладкого тетануса.
Амплитуда гладкого тетануса значительно больше, чем амплитуда одиночного мышечного сокращения .
Если каждый последующий импульс попадает в период расслабления мышцы, то развивается зубчатый тетанус.
Различают следующие фазы одиночного мышечного сокращения.
1.Латентный период.
2.Период ускорения.
3.Период расслабления
40. нарисуйте миограмму одиночного мышечного сокращения и обозначьте его фазы.
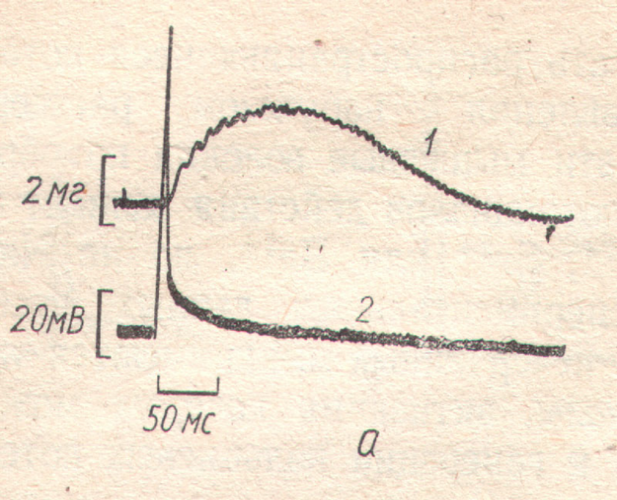
Различают следующие фазы мышечного сокращения.
1.Латентный период.
2.Период ускорения.
3.Период расслабления.
1. Латентный период - это время от момента нанесения раздражения до начала сокращения. Он равен 1,5 мсек.
Структура латентного периода:
"Латентное расслабление", 2) развитие потенциала действия, 3) увеличение объема, т.к.увеличивается концентрация ионов, которые связывают воду, 4) увеличение рН, 5) уменьшение емкостноомического сопротивления, 6) возникает тепло активации.

41. что такое латентный период одиночного мышечного сокращения?
1. Латентный период - это время от момента нанесения раздражения до начала сокращения. Он равен 1,5 мсек.
Структура латентного периода:
"Латентное расслабление", 2) развитие потенциала действия, 3) увеличение объема, т.к.увеличивается концентрация ионов, которые связывают воду, 4) увеличение рН, 5) уменьшение емкостноомического сопротивления, 6) возникает тепло активации.
42. что такое тетаническое сокращение? назовите виды тетануса.
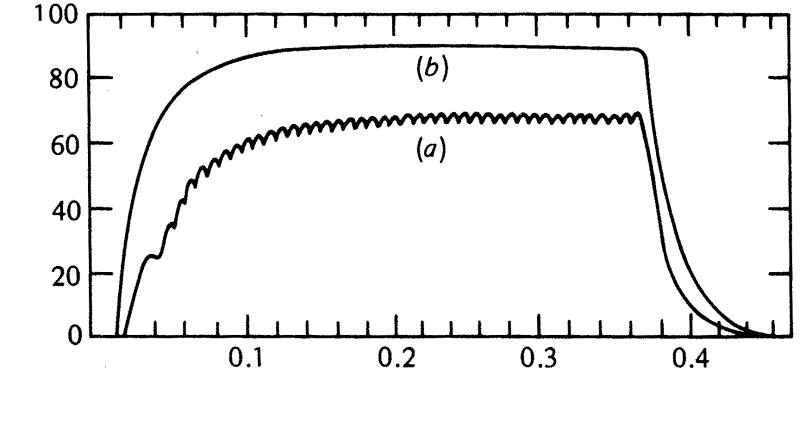
Мышца может быть возбуждена еще во время сокращения - при ритмическом раздражении.
Тетанус - слитное сокращение, возникающее при ритмической стимуляции, когда время между сокращениями меньше, чем длительность одиночного сокращения. В связи с тем, что во время сокращения мышца не имеет рефрактерности, повторные стимулы, наносимые до расслабления, способны создавать дополнительную активацию сократительных элементов и увеличивать силу мышечного сокращения. Этот феномен носит название гладкого тетануса.
Амплитуда гладкого тетануса значительно больше, чем амплитуда одиночного мышечного сокращения .
Если каждый последующий импульс попадает в период расслабления мышцы, то развивается зубчатый тетанус.
Механизм тетануса
П.Е. Введенский считал, что увеличение амплитуды тетанического сокращения развивается вследствие посттетанической потенциации мышц. Т.е. предыдущее сокращение влияет на эффект последующего вследствие фазного изменения возбудимости в связи с развитием потенциала действия. Следовательно, величина тетанического сокращения строго зависит от того, в какую фазу изменения возбудимости после действия предыдущего раздражения приходится действие последующего раздражителя.
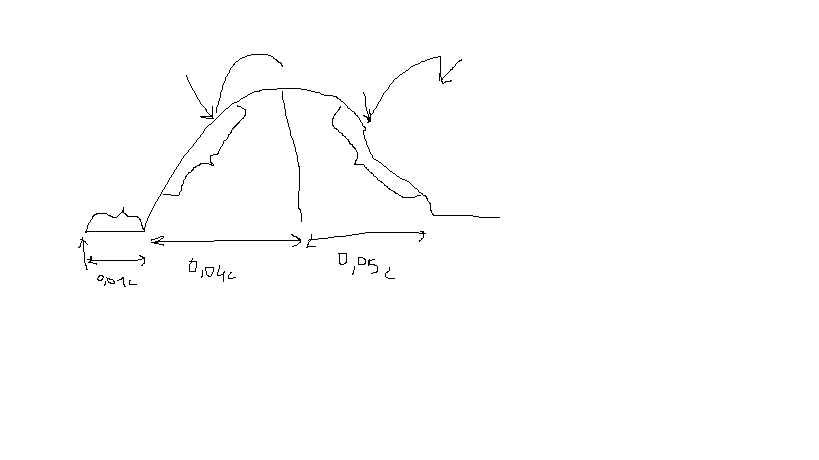
43. в чем состоят особенности тетануса в естественных условиях?
В мышце возникает пространственная суммация, т.е. естественное сокращение возникает посредством суммации одиночных сокращений множества волокон, причем каждое волокно в естественных условиях сокращается много раз. Однако, благодаря значительной инерции мышцы в целом эти отдельные сокращения сливаются в единое сокращение. Этому способствует то, что мотонейроны посылают свои импульсы к мышце не одновременно, и следовательно, в каждый момент времени сокращаются различные группы мышечных волокон.
44. что такое нейромоторная единица? Классификация нейромоторных единиц.
Это совокупность мышечных волокон, иннервируемых одним и тем же мотонейроном.
Фазные быстрые НМЕ имеют мало терминалей и соответственно мало иннервируемых мышечных волокон. В миофибриллах хорошо развит сарко-плазматический ретикулум (СПР), Т-система, а также активен кальциевый насос. Эти НМЕ делятся на НМЕ гликолитического и окислительного типов.
У НМЕ гликолитического типа нет миоглобина, поэтому они имеют белый цвет. У них мало митохондрий, АТФ образуется в основном по механизму гликолиза и быстро расщепляется, т.к. в этих НМЕ высокоактивна АТФ-аза. Они сокращаются быстро, сильно и сразу утомляются. Таких НМЕ много в мышцах гортани (НМЕ включает 2-3 миофибриллы) и в мышцах глазного яблока (НМЕ включает 3-6 миофибриллы).
Фазные быстрые НМЕ окислительного типа занимают промежуточное положение между фазными быстрыми НМЕ гликолитического типа и фазными медленными НМЕ. В них содержится достаточно много миоглобина, имеется много митохондрий, синтезируется достаточное количество АТФ по механизму окислительного фосфорилирования. Однако активность АТФ-азы у них низкая. Они сокращаются быстро, сильно и достаточно долго. Такие НМЕ преобладают в мышцах длительно летающих птиц.
Фазные медленные НМЕ получают иннервацию от α-мотонейронов, порог раздражения которых меньше чем у таковых нейронов, иннервирующих быстрые НМЕ. Аксоны этих нейронов имеют много терминалей и иннервируют соответственно много миофибрилл (от сотен до 2000). У них хуже, чем у быстрых НМЕ, развиты СПР и Т-система. У них много миоглобина, поэтому они имеют красный цвет, много митохондрий, АТФ образуется по механизму окислительного фосфорилирования, а активность АТФ-азы низкая. Эти НМЕ сокращаются медленно, сильно и долго без утомления, а после утомления быстро восстанавливаются. Такие НМЕ преобладают в мышцах, поддерживающих позу человека, четырёхглавой мышце бедра, икроножной и дыхательных мышцах.
Тонические НМЕ сокращаются очень медленно слабо и долго (собственно поэтому они называются тоническими). У человека они имеются только в наружных мышцах глаз, но широко представлены в мышцах амфибий и рептилий. Они иннервированы γ-мотонейронами, имеют много терминалей и соответственно образуют синапсы на множестве миофибрилл. На их мембранах нет потенциалзависимых натриевых каналов и поэтому они генерируют только местное возбуждение.
45. назовите режимы сокращения скелетных мышц.
Режимы сокращения
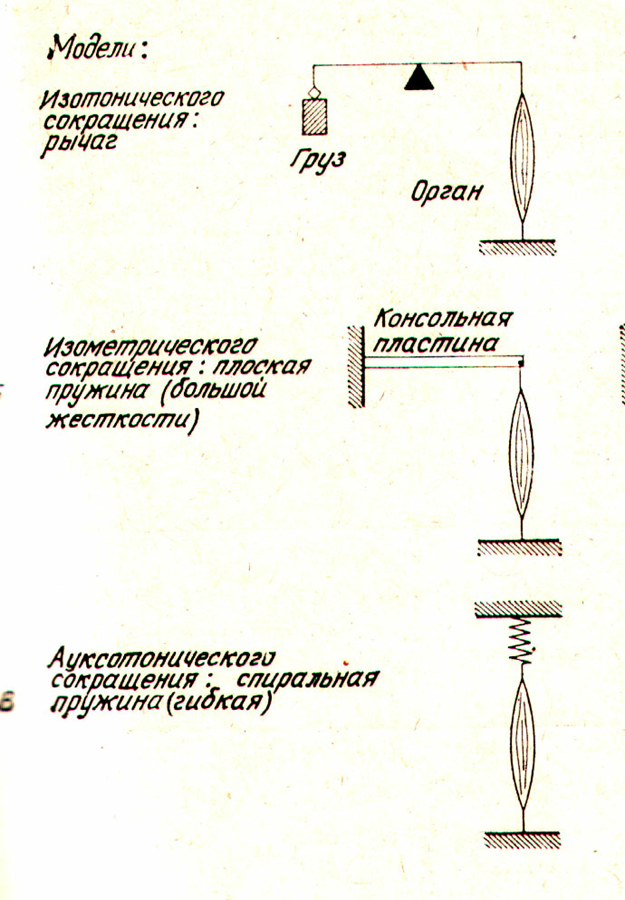
В зависимости от условий, в которых происходит мышечное сокращение, различают два его типа - зотоническое и изометрическое.
Изотоническим называется такое сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным.
Изометрическим называют такое сокращение, при котором мышца укорачиваться не может, т.е. когда оба ее конца неподвижно закреплены. В этом случае длина мышечные волокон остается неизменной, а напряжение их по мере развития сократительного процесса возрастает.
Ауксотоническое сокращение - эти условия создаются когда мышца развивает силу пропорциональную перемещению.
46. что является структурно-функциональной единицей миофибрилл? Опишите ее строение.
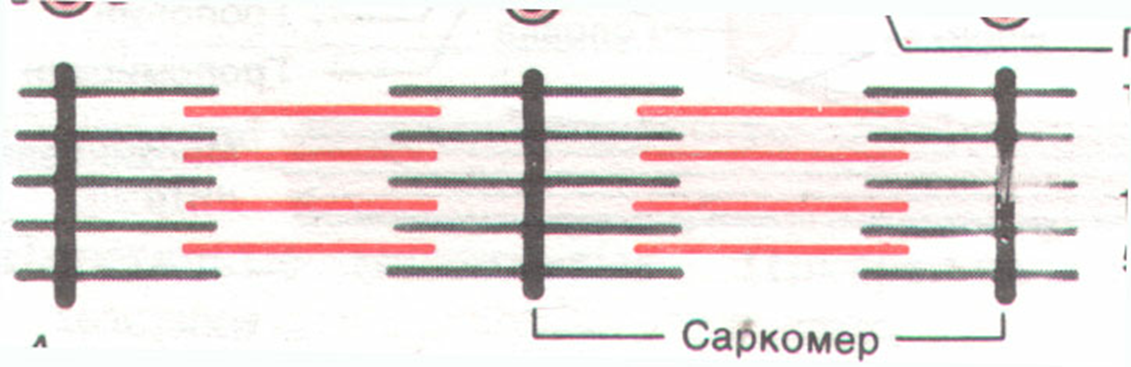
Расстояние от одной Z-пластинки до другой составляет 2,5 мкм и называется саркомером
Сарколемма - это клеточная мембрана мышечного волокна. Однако сарколемма состоит из обычной клеточной мембраны снаружи покрытой тонким слоем полисахаридов и коллагеновых нитей. На конце мышечного волокна этот поверхностный слой сарколеммы склеивается с сухожильным волокном, а сухожильные волокна в свою очередь собираются в пучки, формируя мышечные сухожилия, которые прикрепляются к костям. Сарколемма образует инвагинации, которые увеличивают площадь поверхности мембраны (Т-трубочки).
47. опишите электронномикроскопическую картину миофибрилл.
Сарколемма - это клеточная мембрана мышечного волокна. Однако сарколемма состоит из обычной клеточной мембраны снаружи покрытой тонким слоем полисахаридов и коллагеновых нитей. На конце мышечного волокна этот поверхностный слой сарколеммы склеивается с сухожильным волокном, а сухожильные волокна в свою очередь собираются в пучки, формируя мышечные сухожилия, которые прикрепляются к костям. Сарколемма образует инвагинации, которые увеличивают площадь поверхности мембраны (Т-трубочки
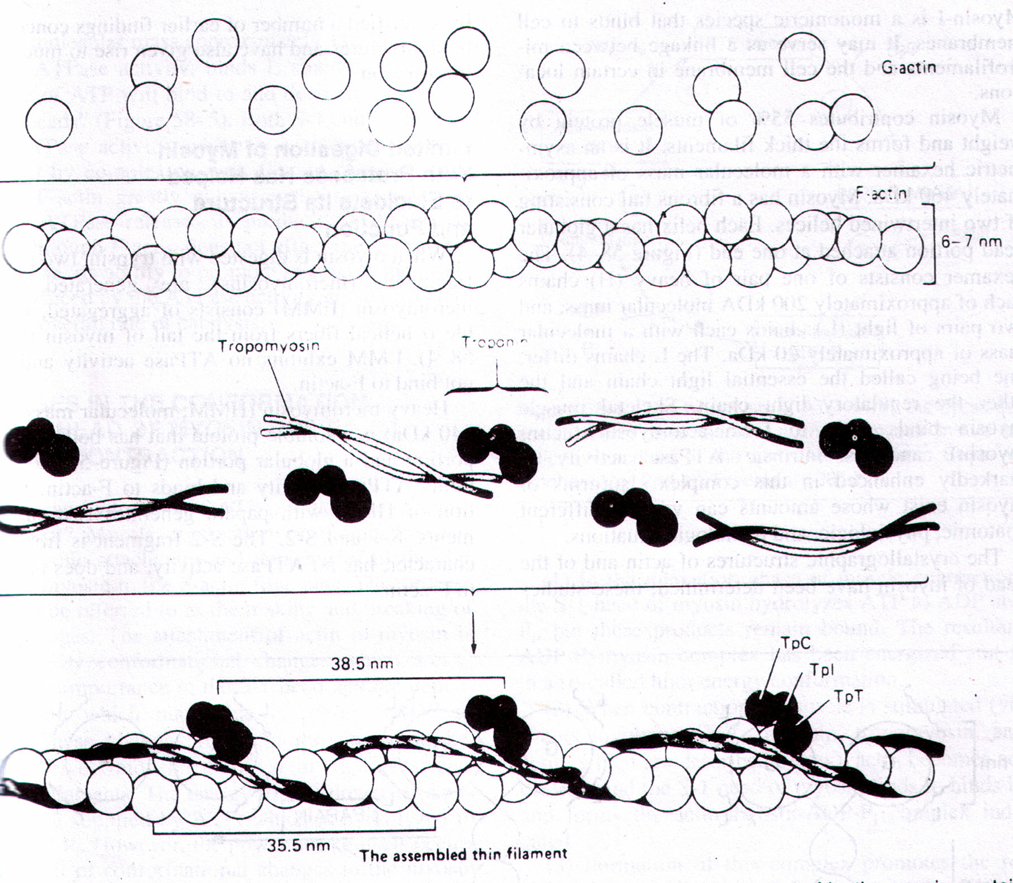
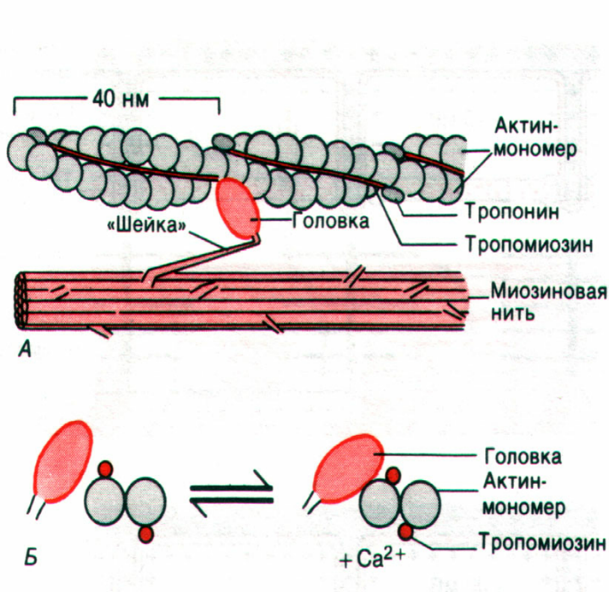
Актиновый миофиламент представляет собой 400 молекул сократительного белка актина в виде тонких двойных нитей, закрученных в двойную спираль с шагом 36,5 нм. Длина актинового миофиламента составляет примерно 1 мкм, а диаметр – 5 нм. В нём имеются активные центры, располагающиеся друг от друга на расстоянии 20 нм. Молекулярная масса белка актина 42000 дальтон. В бороздках актиновых миофиламентов располагаются молекулы регуляторных белков (эти белки не участвуют прямо в сокращении, но регулируют его) – это тропомиозин и тропонин.
Тропомиозин имеет нитевидную форму и к нему прикрепляется тропонин, имеющий глобулярную форму.
Тропонин имеет три субъединицы:
1) TN-C – это кальцийсвязывающая субъединица;
2) TN-I – это ингибирующая субъединица, она после связывания TN-C с кальцием изменяет свою конформацию и тропомиозин идёт вглубь (в желобок) спирали, открывая при этом активные центры актинового миофиламента;
3) TN-T – это субъединица, связывающая тропонин с тропомиозином.
Миофибриллыг. Актиновые и миозиновые нити. Каждое мышечное волокно содержит от нескольких сотен до нескольких тысяч миофибрилл. Каждая миофибрилла состоит приблизительно из 1500 миозиновых нитей и 3000 актиновых нитей. Миозиновые и актиновые нити частично заходят друг за друга, что и определяет двойное лучепреломление в темном диске.Светлые диски содержат только актиновые нити (диски). Миозиновая нить является более толстой, чем актиновая. По сторонам миозиновой нити выступают маленькие шипики, которые называются попереч¬ными мостиками. Концы актиновых нитей прикрепляются к так назывемым дискам. От этих дисков нити идут в обе стороны, заходя между миозиновыми нитями.
Таким образом, мышечное волокно состоит из светлые и темным дисков. Это придает мышце исчерченность.
Часть миофибрилл, которая лежит между двумя мембранами называется саркомером.
Когда скелетная мышца полностью расслаблена и растянута длина саркомера равна 2 мкм. При этой длине нити актина полностью перекрывают нити миозина и даже немного перекрываются между собой.
Саркоплазма- это внутренняя среда мышечной клетки. В ней содержится много митохондрий, которые лежат параллельно миофибриллам.
Саркоплазматический ретикулум. В саркоплазме накалится многочисленный Саркоплазматический ретикулум (СПР). Ретикулум имеет специальную организацию, которая чрезвычайно важна в контроле мышечного сокращения,которое обсуждается в следу¬ющей главе. Чем быстрее способна сокращаться мышца , тем более развитый СПР она имеет.
В СПР различают цистерны СПР и продольные трубочки. Цистерны СПР близко подходят к поперечным трубочкам (Т-тру-бочкам) образуя триаду ( цистерна, Т- трубочка и цистерна).
48. опишите строение тонкого миофиламента.
Актиновый миофиламент представляет собой 400 молекул сократительного белка актина в виде тонких двойных нитей, закрученных в двойную спираль с шагом 36,5 нм. Длина актинового миофиламента составляет примерно 1 мкм, а диаметр – 5 нм. В нём имеются активные центры, располагающиеся друг от друга на расстоянии 20 нм. Молекулярная масса белка актина 42000 дальтон. В бороздках актиновых миофиламентов располагаются молекулы регуляторных белков (эти белки не участвуют прямо в сокращении, но регулируют его) – это тропомиозин и тропонин.
Тропомиозин имеет нитевидную форму и к нему прикрепляется тропонин, имеющий глобулярную форму.
Тропонин имеет три субъединицы:
1) TN-C – это кальцийсвязывающая субъединица;
2) TN-I – это ингибирующая субъединица, она после связывания TN-C с кальцием изменяет свою конформацию и тропомиозин идёт вглубь (в желобок) спирали, открывая при этом активные центры актинового миофиламента;
3) TN-T – это субъединица, связывающая тропонин с тропомиозином
49. что представляет собой толстый миофиламент?
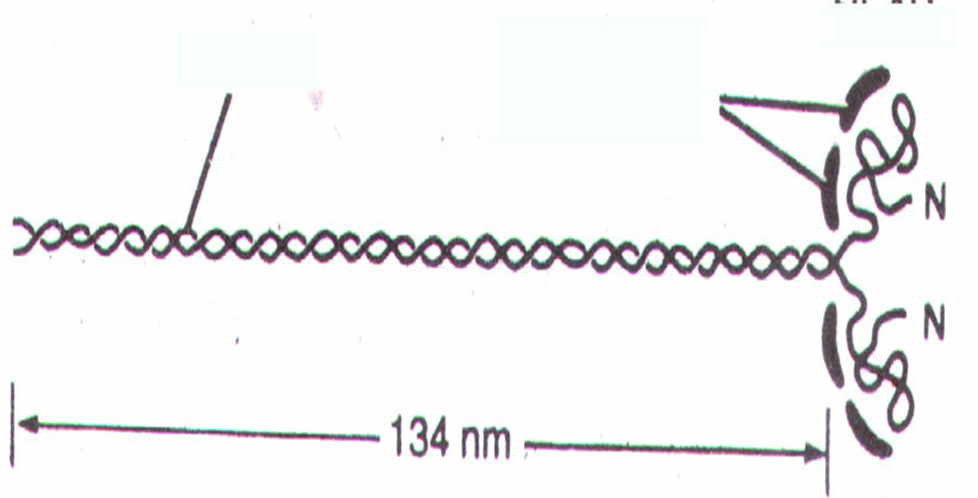
Миозиновый миофиламент имеет длину примерно 1,6 мкм и диаметр – 10 нм и состоит примерно из 300 молекул белка миозина (молекулярная масса 500000 дальтон). Молекула миозина парная, имеет две головки, две шейки и два хвоста. В головке миозина имеется 2 субфрагмента: S1 – тяжёлые цепи; S2 – лёгкие цепи, которые продолжаются в шейку. Шейка миозина эластичная, представляет собой шарнирное соединение и головка может поворачиваться на шейке вокруг своей оси. В хвосте миозина находятся лёгкие цепи, молекулы миозина соединяются между собой хвостами. На боковых сторонах миозинового филамента имеются выступы, которые называются поперечными мостиками, они ориентированы к оси миозиновой нити под углом 120о. Поперечный мостик состоит из головки и шейки миозиновой молекулы.
Функция головок миозиновой молекулы!
Осуществляется связь миозина с актином
Способна гидролизовать АТФ.
Предполагают, что роль активного центра на молекуле актина играет молекула АДФ
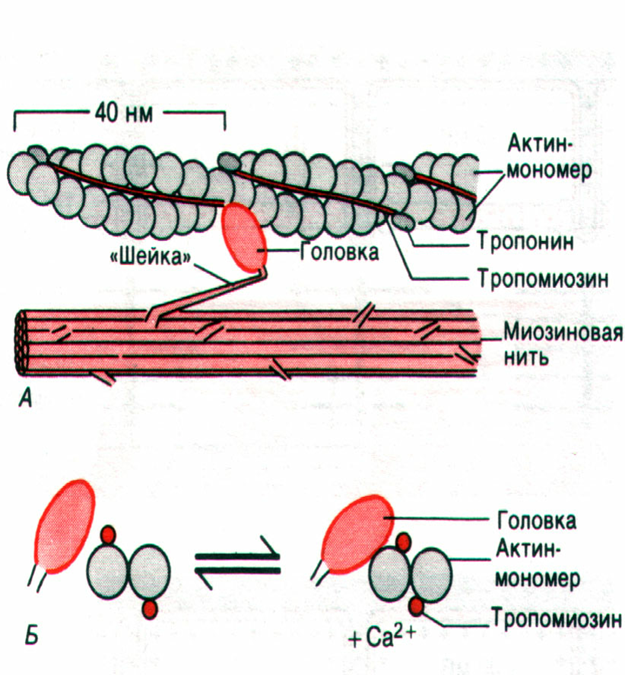
50. какова структура молекул миозина?
Молекулы миозина имеют молекулярную массу около 5000 тысяч и на электронных микрофотографиях выглядят как длинные палочковидные структуры с двумя глобулярными головками.
Большинство известных к настоящему времени миозинов ( II типа ) имеют в общем сходную структуру. Это гексамеры, состоящие из двух больших субъединиц с молекулярной массой около 200 кДа ( тяжелые цепи миозина ) и четырех малых субъединиц с молекулярной массой около 20 кДа ( легкие цепи миозина ). Таким образом, молекулярная масса всей миозиновой молекулы составляет 450 - 500 кДа.

Тяжелые цепи на большом протяжении закручены в двойную спираль и образуют стержневую часть молекулы. На N-конце каждая тяжелая цепь образует глобулярную головку грушевидной формы. Стержневая часть миозина вследствие высокого содержания альфа-спиралей является прямой и жесткой, изгибаться она может только в двух участках. Такие участки, не содержащие альфа-спиралей, называют "шарнирными". Один из них, расположенный в стержневой части миозина, дает ей возможность изгибаться в этой области, а другой участок обеспечивает высокую подвижность головок. Легкие цепи ( LC ) ассоциированы с головками миозина, по 2 с каждой головкой.
51. опишите строение тропонина и укажите роль в инициации мышечного сокращения.
Тропомиозиновые молекулы
Актин также содержит молекулу тропомиозина. Каждая молекула тропомиозина имеет М.в. 70000 и длину 40 нм. Молекула тропомиозина закручена по спирали вдоль борозды F-актина. Считается, что в состоянии покоя молекула тропомиозина лежит на активных центрах актина, экранируя их. Это препятствует взаимодействию актина и миозина. Каждая молекула тропомиози¬на закрывает приблизительно 7 из этих актиновых центров.
Тропонин и его роль в инициации мышечного сокращения.
По ходу спирали F-актина через равные промежутки прикреплены молекулы тропонина. Тропонин - это комплекс, состоящий из 3 белковых молекул слабо скрепленных одна с другой. Первая субьединица тропонина получила название тропонин-I. Эта субьединица обладает высокой аффинностью к актину. Другая субьединица (тропонин Т) обладает высокой аффинностью к тропомиозину. Третья субьединица (тропонин С) обладает сродством к ионам кальция.
52. где расположен в мышечной клетке кальциевый насос? Каково его значение?
Са является одним из наиболее биологически важных элементов, участвующем в обмене веществ различных органов и тканей животных весьма многообразно. Он может поглощаться клеткой пассивно, проходя через мембрану или по градиенту концентраций, или по электрическому полю, или обмениваться на ионы натрия. Однако некоторые функции организма обеспечиваются только энергозатратным переносом Са2+, например, сокращение мышечных волокон, где Са2+ выступает в роли вторичного мессенджера.
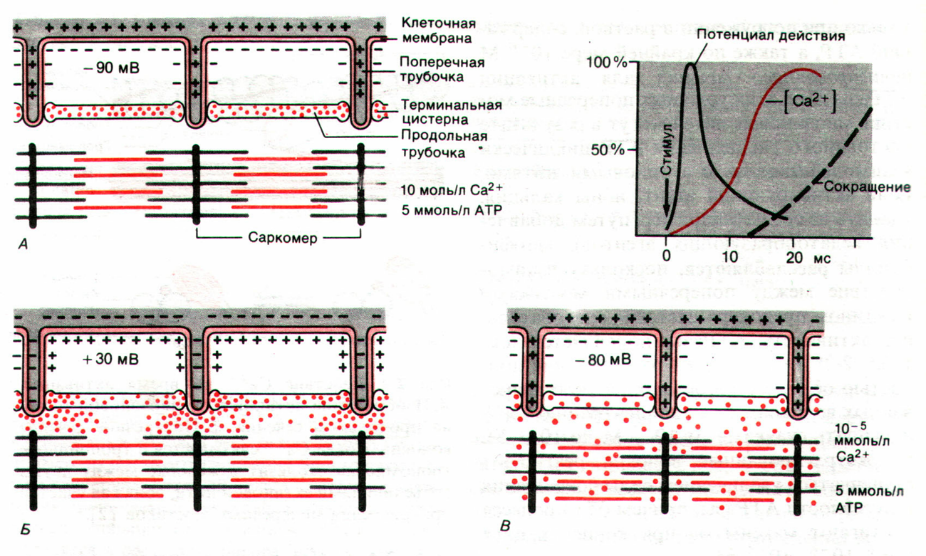
Для понимания работы кальциевого насоса необходимо кратко рассмотреть строение мышечного волокна и механизм его сокращения. Белки, вытянутые от одного края мышечного волокна до другого, представляют собой две сократительные нити — актин и миозин. При сокращении мышцы они проскальзывают друг в друга, а при расслаблении возвращаются в исходное положение. Мышечное волокно насквозь пронизано мембранной сетью, называемой саркоплазматическим ретикулумом (CP). Сократительный элемент (саркомер) состоит из многих нитей актомиозина и CP, причем последний выглядит, как впяченная внутрь волокна клеточная мембрана, опоясывающая сократительный элемент.
В покоящихся клетках миофибрилл концентрация Са2+ мала (ниже 10‑5 М), тогда как в CP она существенно выше (10‑3i М). Высокая концентрация в CP обеспечивается Са2+-АТФазами и поддерживается с помощью специального кислого белка калъсеквестрина (55 кДа). Потенциал действия, поступающий с концевой пластинки двигательного нейрона, деполяризует плазматическую мембрану через поперечные трубочки Т-системы, которые представляют собой трубчатые впячивания клеточной мембраны и тесно контактируют с миофибриллами. В результате потенциал-управляемый мембранный белок ("SR-foot") прилегающей мембраны СРz открывает Са2+-каналы для выброса Са2+ в пространство между филаментами актина и миозина до уровня >10‑5 М. Этот выброс запускает процесс сокращения миофибрилл (рис. 5).
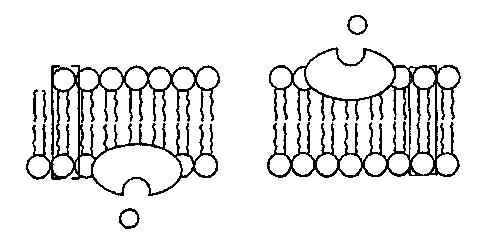
Механизм этого процесса далее представляется следующим образом: выброшенный Са2+ связывается с С-субъединицей тропонина, перестраивая его структуру. Тропонин-миозиновый комплекс разрушается и освобождает на молекуле актина участок связывания с миозином, что и начинает цикл сокращения. После него уровень Са2+ снижается из-за активного обратного переноса в CP, тропонин С отдает Са2+, комплекс тропонин-тропомиозин занимает на молекуле актина исходное положение, блокируя актин-миозиновый цикл. Наступает расслабление мышцы. В клетках эукариот Са2+ может связываться не только с тропонином С, но также с близкими по свойствам кальмодулином и парвальбумином. Такой комплекс непосредственно связан с Са-насосом.
Поскольку высокая концентрация Са2+ в цитоплазме из-за цитотоксического действия иона нежелательна, она быстро снижается благодаря очень активным многочисленным транспортным Са-КТФазам. Химически они представляют собой протеолипиды. По расчетам, молекулы кальциевого насоса занимают треть поверхности мембран мышечной ткани. При гидролизе одной молекулы АТФ внутрь пузырьков CP транспортируется 2 иона Са2+. Как и у натриевого насоса, здесь активный центр связывается с АТФ и 2 Са2+ на мембране со стороны цитозоля, затем он поворачивается внутрь пузырька, выбрасывает Са2+ и АДФ, после чего принимает исходное положение. Таким образом, мышечное сокращение — это механическая энергозатратная работа, обеспечиваемая гидролизом АТФ. Катализирует гидролиз АТФ сам миозин, причем в отсутствие Са2+ распад АТФ и сокращение миофибрилл полностью исчезают. Вероятно, места связывания ШТФ и Са2+ на миозине взаимно конкурентны.
Многочисленные исследования показали, что повышенное содержание Са2+ внутри клетки предшествует развитию программируемой смерти клетки (апоптозу). Часто снижение количества Са2+ отдаляет начало апоптоза. На инициирование этого процесса влияет также содержание Zn2+, действующего по отношению к Са2+ антагонистически.
Влияние этих катионов сказывается прежде всего на изменении мембран клеток, подвергающихся апоптозу. При апоптозе активируются два вида ферментов — эндонуклеаза, расщепляющая ДНК во внутренних участках нуклеосом, и тканевая трансглутаминаза, ковалентно связывающая белки с мембраной посредством образования изопептидных связей. Zn2+ блокирует апоптоз по механизму подавления активности эндонуклеазы. Кроме того, эти катионы влияют на активность антиокислительной системы путем взаимодействия с металлами, координирующими ферменты этой системы, прежде всего с Fe и Си.
В ходе эволюции переход от водной среды с большим содержанием Са2+ к наземному существованию был сопряжён с развитием сложного механизма гомеостаза иона, поскольку он цитотоксичен, для предотвращения резких изменений концентрации Са2+ во внеклеточной жидкости. В этом механизме основную роль играют три гормона — паратиреоидный (ПТГ), кальцитриол (витамин Д) и кальцитонин (КТ). Механизм гомеостаза кальция очень эффективен, а при сбое в его работе наблюдаются разнообразные патологические состояния, в том числе резко сокращается продолжительность жизни организма.
Общим для натриевого и кальциевого насосов является свойство всегда переносить положительные заряды асимметрично из клетки во внеклеточную среду, только у натриевого насоса этой средой является внешняя среда, а у кальциевого — специализированные компартменты (пузырьки CP) внутри клетки. Таким образом, кальциевый насос является специализированной системой, предназначенной для быстрой регуляции концентрации Са2+ в скелетных мышцах.
53. в чем состоит теория «скольжения», объясняющая механизм мышечного сокращения?
теория скольжения
Далее происходит присоединение головки поперечного мостика миозинового филамента к активным центрам актинового филамента. При этом головка миозина поворачивается вокруг своей оси, т.к. имеет несколько активных центров, которые последовательно взаимодействуют с активными участками на актиновом филаменте, развивая при этом тянущие усилия.
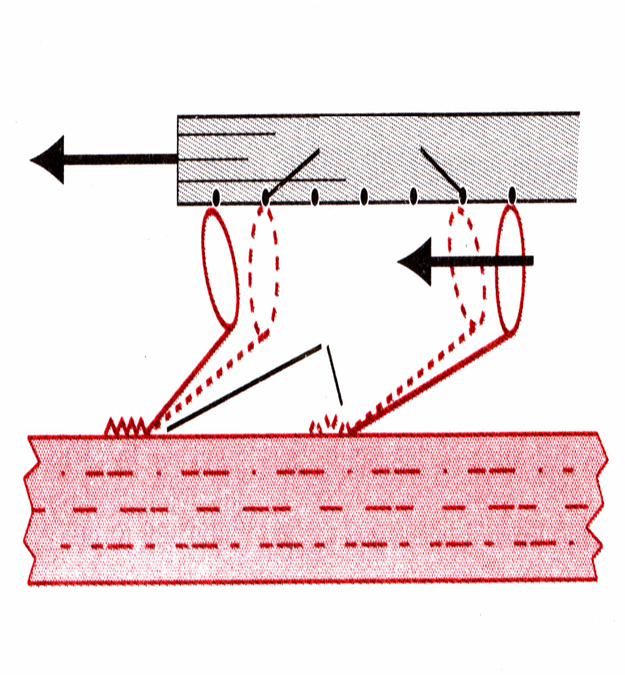
ТЕОРИЯ СКОЛЬЖЕНИЯ
После того как головки миозиновых молекул присоединяются к активным центрам
актина, одновременно происходят изменения во внутримолекулярные силах между головкой миозина и субфрагментом 1. Головка в области шарнирного механизма сгибается по направлению к 2 и тем самым проталкивает актиновую молекулу вдоль миозиновой молекулы. Этот наклон головки называется (силовой удар). После этого наклона головка миозино¬вой молекулы автоматически отсоединяется от активного центра на молекуле актина. Затем головка возвращается в нормальное положение и в этомположении вновь соединяется с новым актив¬ным центром, вновь отсоединяется, а актиновая нить продвигается еще на один шаг.
Считается, что головка миозиновой молекулы работает независима от других головок.
54. в чем заключается кризис теории «скольжения»?
В настоящее время появились некоторые факты, которые не вписываются в классическую теорию скольжения:
1) миозиновый филамент при сокращении меняет свою длину и диаметр (утолщается и укорачивается);
2) при сокращении укорачивается не только диск I, но и диск А;
3) между актиновыми и миозиновыми филаментами расстояние – 13 нм, а длина головки миозина – 19-21 нм;
4) сокращение мышцы ступенчатое, а не плавное, как при скольжении.
55. что такое минимальный сократительный элемент?
На основании теории вкручивания в качестве структуры сократительного аппарата выделяют минимальный.сократительный элемент (МСЭ). МСЭ представляет собой миозиновый филамент, окруженный шестью актиновыми филаментами.
Анализ строения МСЭ показывает, что его центральная часть - миозиновый филамент напоминает своеобразный винт. Особенность строения винта состоит в том, что трехзаходная спираль, образованная головками молекул миозина, образует широкую, практически ленточную однозаходную спираль. Определенные особенности наблюдаются также и в структуре актиновых филаментов в связи с наличием тропонин-тропомиозинового комплекса. Глобулы тропонина С попадают в широкую бороздку, образованную головками молекул миозина в структуре толстого филамента. Такое взаимное расположение структур в МСЭ делает практически невозможным взаимное скольжение актиновых и миозиновых нитей.
56. в чем состоит теория «вкручивания», объясняющая механизм мышечного сокращения?
Авторы считают, что в основе сокращения лежит вкручивание миозиновых филаментов в трубкообразные структуры, образованные актиновыми филаментами, а само сокращение обеспечивается междоменными перемещениями в работающих по очереди головках миозина
На основании этой теории в качестве структуры сократительного аппарата выделяют минимальный.сократительный элемент (МСЭ). МСЭ представляет собой миозиновый филамент, окруженный шестью актиновыми филаментами.
Анализ строения МСЭ показывает, что его центральная часть - миозиновый филамент напоминает своеобразный винт. Особенность строения винта состоит в том, что трехзаходная спираль, образованная головками молекул миозина, образует широкую, практически ленточную однозаходную спираль. Определенные особенности наблюдаются также и в структуре актиновых филаментов в связи с наличием тропонин-тропомиозинового комплекса. Глобулы тропонина С попадают в широкую бороздку, образованную головками молекул миозина в структуре толстого филамента. Такое взаимное расположение структур в МСЭ делает практически невозможным взаимное скольжение актиновых и миозиновых нитей.
Учитывая все выше сказанное стало очевидным, что перемещение толстых и тонких нитей возможно за счет вкручивания миозиновых филаментов в трубкообразные структуры, образованные актиновыми филаментами. Поскольку код спирали на разных концах миозиновой нити имеет разное направление, то ее закручивание проходит в разные стороны. Аналогичное закручивание можно наблюдать, если концы резиновой трубки вращать в противоположные стороны
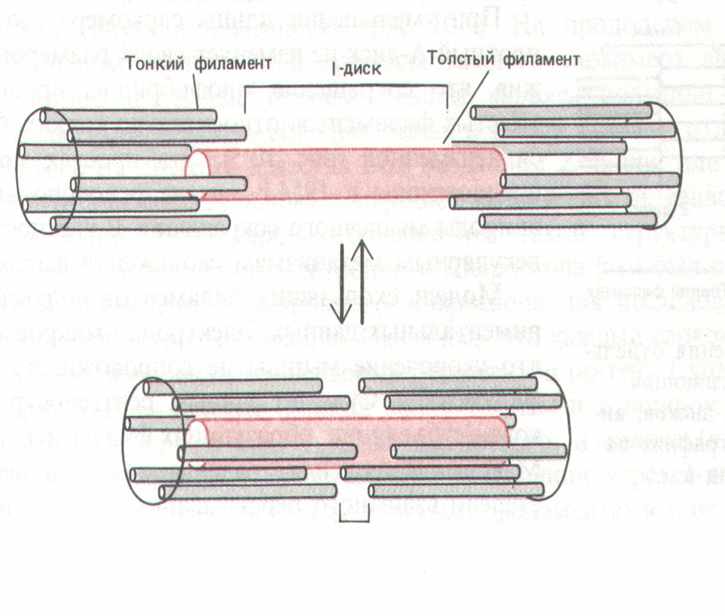
Минимальный
сократительный элемент
Схематично это можно представить следующим образом. При зацеплении головки миозиновой нити за актин, она расположена по отношению к остову миозиновой нити под углом 45о. При сокращении мышцы происходит уменьшение расстояния между зацеплением головки за тонкую нить и ее фиксацией в структуре миозинового филамента, что сопровождается перемещением толстых и тонких филаментов относительно друг друга. Головка в конце рабочего хода занимает практически перпендикулярное положение по отношению к филаментам. При закручивании нити миозина головки работают по-очереди, вначале присоединяется одна головка миозиновой молекулы и подтягивает актиновую нить, что дает возможность другой миозиновой головке присоединиться к активному центру на другой молекуле актина и т.д. В результате миозиновая нить закручивается, укорачивается, что и приводит к уменьшению длины А-диска.
57. опишите компоненты проводящей системы мышечного волокна (мембранно- миофибриллярная связь).
Осуществляет проводящая система мышечного волокна(триада):
1.цитоплазматическая мембрана мышечного волокна.
2. Т-трубочка – инвагинация цитоплазматической мембраны мышечного волокна
3. l-трубочка – терминальная цистерна саркоплазматического ретикулума (СПР). Мембраны l- и Т-трубочек непосредственно соединены посредством соединительных ножек.
Спр содержит ионы кальция в концентрации 10-4 моль/л. Их высвобождение происходит в тот момент, когда Т-трубочки приходят в состояние возбуждения. Благодаря тому, что концентрация кальция в межмиофибриллярном пространстве 10-8 моль/л, его выброс происходит мгновенно – «кальциевый залп». Насосы, расположенные в стенке спр, возвращают ионы кальция из цитоплазмы, в результате чего их концентрация снижается до уровня ниже того, при котором может быть инициировано сокращение.
Т-трубочка
58. в чем состоит механизм сопряжения возбуждения и сокращения в мышце? Каковы его особенности в скелетной, гладкой и сердечной мышцах?
Сопряжение возбуждения и сокращения в скелетной мышце
В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий.

Рис. 3. Механизм сопряжения возбуждения и сокращения.
Объяснение – в тексте.
1 – поперечная трубочка саркоплазматичекой мембраны,
2 –саркоплазматичекий ретикулум,
3 – ион Са++ ,
4 – молекула тропонина,
5 – молекула тропомиозина.
• В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану.
• Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В).
• Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г).
• К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д).
Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения .
Рассмотрим процесс трансформации потенциала действия в сокращение кардиомиоцитов или процесс сопряжения возбуждения и сокращения. В его основе лежит переход химической энергии в виде макроэргических фосфатов в механическую энергию сокращений кардиомиоцитов.
Существует несколько белков, ответственных за сокращение клеток миокарда (рис. 1.17). Два из них — актин и миозин — являются главными сократительными элементами. Два других — тропомиозин и тропо-нин — выполняют регуляторную функцию.
Миозин образован толстыми филаментами, каждый филамент состоит примерно из 300 продольно расположенных молекул. Филамент миозина имеет шаровидные головки, которые равномерно распределены по его длине и содержат миозиновую АТФ-азу — фермент, необходимый для сокращения. Актин, более мелкая молекула, представлена тонкими филаментами, образующими альфа-спираль, две цепи которой расположены между толстыми филаментами миозина (рис. 1.8). Титин (также называется коннектин), недавно обнаруженный белок, способствует прикреплению миозина к Z-линии саркомера, а также влияет на эластичность сокращения.
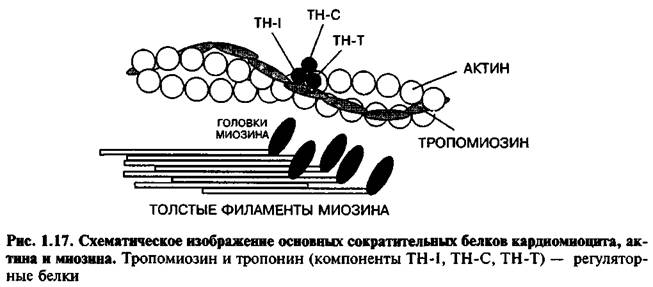
Тропомиозин — двойная спираль, которая лежит в углублении между филаментами актина и в состоянии покоя препятствует взаимодействию между миозиновыми головками и актином, предупреждая тем самым сокращение. Тропонин равномерно распределен вдоль актиновых нитей и состоит из трех субъединиц. Субъединица тропонин С (ТН-С) ответственна за связывание ионов кальция и тем самым — за процесс сокращения. Тропонин I (TH-I) — субъединица, тормозящая активность АТФ-азы, осуществляющую связь актин-миозин. Тропонин Т (ТН-Т) — субъединица, связывающая тропониновый комплекс с актином и молекулами тро-помиозина.
Во время фазы 2 потенциала действия, Са++ проникает в миоциты через кальциевые каналы сарколеммы и Т-каналы. Относительно малое количество кальция, поступившего в клетку, не способно вызвать сокращение миофибрилл, однако оно стимулирует массивный выход Са++ из сарко-плазматического ретикулума (рис. 1.18). В результате этого концентрация кальция в клетке увеличивается в 10 раз.
Когда ионы кальция связаны с ТН-С, тормозится активность ТН-Т; это обусловливает изменение строения тропомиозина. В результате становится возможным взаимодействие между актином и миозином, приводящее к сокращению.
Мышечное сокращение развивается вследствие связывания головок миозина с актиновыми филаментами и сгибания головок. В результате этого тонкие и толстые филаменты движутся друг вдоль друга за счет энергии АТФ (рис. 1.19). Первым этапом в этом процессе является активация головки миозина при гидролизе АТФ, после чего головка миозина связывается с актином, образуя поперечный мостик. Взаимодействие головки миозина с актином приводит к структурным изменениям в головке, вызывающим ее сгибание Это сгибательное движение вызывает смещение актинового филамента вдоль миозинового.
Пока головка миозина все еще связана с актином, дополнительная молекула АТФ связывается с головкой миозина и вытесняет АДФ, образовавшийся при предшествующем гидролизе АТФ. Связывание нового АТФ с головкой миозина приводит к высвобождению филамента актина, а гидролиз АТФ возвращает головку миозина в разогнутое состояние, подготавливая ее к следующему циклу.
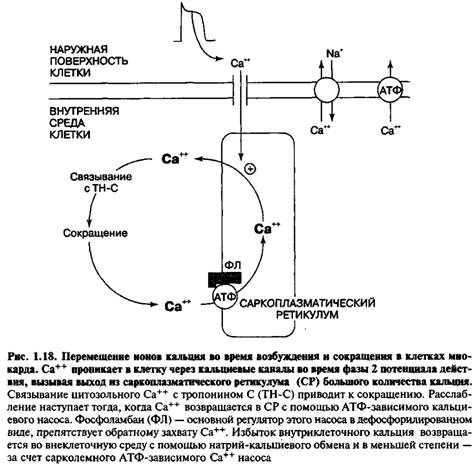
Образование и разрыв связей актина и миозина приводит к укорочению мышечного волокна за счет увеличения степени перекрывания толстых и тонких филаментов внутри каждого сар-комера. В присутствии АТФ этот процесс может продолжаться до тех пор, пока концентрация кальция будет достаточной, чтобы препятствовать блокирующему действию комплекса тропонин-тропомиозин.
В завершение фазы 2 потенциала действия, Са++ каналы закрываются, кальций перестает поступать в саркоплазму. В это время кальций удаляется в саркоплазматический ретикулум или выходит из клетки (рис. 1.18). Разрыв связи ионов кальция с тропонином С приводит к тому, что тропо-миозин блокирует связь актина с миозином, приводя к расслаблению клеток. С началом следующего потенциала действия цикл сокращения и расслабления может вновь повториться.
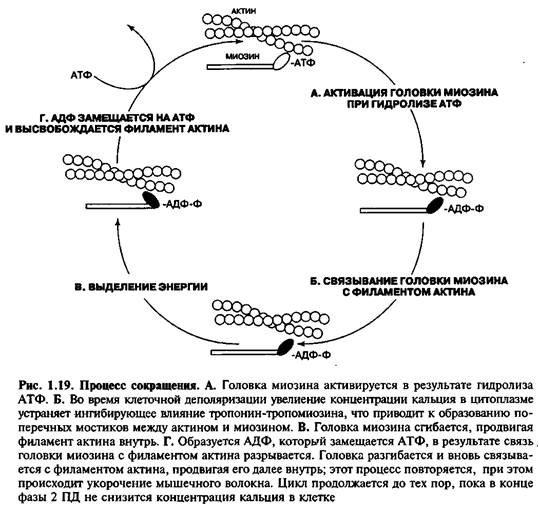
Доказано, что концентрация Са++ внутри клетки является главным фактором, определяющим силу сердечного сокращения. Механизмы, которые способствуют повышению концентрации внутриклеточного кальция, увеличивают силу сокращения, в то время как факторы, снижающие концентрацию кальция — уменьшают силу сокращения.
Роль Са++ в сопряжении возбуждения и сокращения. В отличие от скелетных мышц, где функцию сопрягающего агента между мембраной и актомиозиновым комплексом выполняет Т-система и СПР, в ГМК эти образования либо отсутствуют (Т-система), либо их недостаточно (СПР составляет примерно 2-7% объема цитоплазмы). В ГМК функцию сопряжения выполняют, очевидно, кавеолы плазматической мембраны ГМК и приближенные к ним участки СПР.
Важную роль в сопряжении процесса возбуждения и сокращения играют ионы Са++, которые снимают тормозящее воздействие тропонина на актин-миозиновое взаимодействие и запускают сокращения. В ГМК исходная концентрация Са++ составляет 2,0-2,8´10-7 моль/л, повышаясь до 3,8´10-6 моль/л на максимуме сокращения. Источниками Са++ для ГМК служат: 1) ионизированный Са++ внутриклеточной среды; 2) Са++ токи плазматической мембраны через 2 класса Са++-каналов и Na+-неселективные каналы; 3) внутриклеточные депо Са++ – митохондрии и СПР, внутренняя поверхность плазматической мембраны с ее Са++-связывающими белками. В отличие от скелетной мышцы, в ГМК ведущая роль принадлежит не СПР, а плазмалеммальным токам, т.к. благодаря относительно малому объему каждой клетки по сравнению с поперечно-полосатым мышечным волокном диффузионное расстояние плазмалемма – миофибрилла падает.
59. какова роль кальция в мышечном сокращении?
Доказано, что концентрация Са++ внутри клетки является главным фактором, определяющим силу сердечного сокращения. Механизмы, которые способствуют повышению концентрации внутриклеточного кальция, увеличивают силу сокращения, в то время как факторы, снижающие концентрацию кальция — уменьшают силу сокращения.
Роль Са++ в сопряжении возбуждения и сокращения. В отличие от скелетных мышц, где функцию сопрягающего агента между мембраной и актомиозиновым комплексом выполняет Т-система и СПР, в ГМК эти образования либо отсутствуют (Т-система), либо их недостаточно (СПР составляет примерно 2-7% объема цитоплазмы). В ГМК функцию сопряжения выполняют, очевидно, кавеолы плазматической мембраны ГМК и приближенные к ним участки СПР.
Важную роль в сопряжении процесса возбуждения и сокращения играют ионы Са++, которые снимают тормозящее воздействие тропонина на актин-миозиновое взаимодействие и запускают сокращения. В ГМК исходная концентрация Са++ составляет 2,0-2,8´10-7 моль/л, повышаясь до 3,8´10-6 моль/л на максимуме сокращения. Источниками Са++ для ГМК служат: 1) ионизированный Са++ внутриклеточной среды; 2) Са++ токи плазматической мембраны через 2 класса Са++-каналов и Na+-неселективные каналы; 3) внутриклеточные депо Са++ – митохондрии и СПР, внутренняя поверхность плазматической мембраны с ее Са++-связывающими белками. В отличие от скелетной мышцы, в ГМК ведущая роль принадлежит не СПР, а плазмалеммальным токам, т.к. благодаря относительно малому объему каждой клетки по сравнению с поперечно-полосатым мышечным волокном диффузионное расстояние плазмалемма – миофибрилла падает.
Ключевая роль в регуляции мышечного сокращения принадлежит ионам кальция (Са2+). Миофибриллы обладают способностью взаимодействовать с АТФ и сокращаться лишь при наличии в среде определенных концентраций ионов кальция. В покоящейся мышце концентрация ионов Са2+ поддерживается ниже пороговой величины при участии Са2+-зависимой АТФазы. В состоянии покоя эта система активного транспорта накапливает кальций в цистернах саркоплазматического ретикулума и трубочках Т-системы.
Мышечное сокращение инициируется приходом потенциала действия на концевую пластинку двигательного нерва. В синапс выделяется ацетилхолин, который связывается с постсинаптическими рецепторами мышечного волокна. Далее потенциал действия распространяется вдоль сарколеммы к поперечным трубочкам Т-системы и происходит передача сигнала на цистерны саркоплазматического ретикулума. Последние начинают освобождать находящийся в них кальций в саркоплазму. Концентрация Са2+ увеличивается с 10-7 до 10-5 ммоль/л. Кальций связывается с Тн-С, что вызывает конформационные сдвиги, передающиеся на тропомиозин и далее - на актин. Открываются закрытые ранее центры в актине для связывания с миозином. Актин взаимодействует с миозином, что инициирует сокращение мышечного волокна.
После прекращения действия двигательного импульса кальций с помощью Са2+-зависимой АТФазы откачивается из цитоплазмы в цистерны саркоплазматического ретикулума. Уход кальция из комплекса с Тн-С приводит к смещению тропомиозина и закрытию активных центров актина. Миозиновая «головка» отсоединяется от актина. Мышца расслабляется.
Кальций является аллостерическим модулятором мышечного сокращения.
Деполяризация Т-трубочек
Выброс Са2+ из цистерн
саркоплазматического ретикулума
Комплекс Тн-С + 4Са2+
Тропонин (активный)
Тропомиозин (активный)
F – актин
Актин – Миозин
АДФ, Фн
сократительный цикл
Чистая нить актина в отсутствии тропонин-тропомиозинового комплекса обладает способностью сильно связываться с молекулой миозина в присутствии Мg2+ и АТФ, которые в миофибрилах находятся в достаточном количестве. Однако, если к нитям филамента добавить тропонин-тропомиозиновый комплекс, взаимодействие актина и миозина ингибируется. Поэтому считается, что тропонин-тропомиозиновый комплекс блокирует сокра¬щение и обеспечивает расслабление скелетной мышцы. В присутствии ионов Са2+ его 4 молекулы связываются с каждой молекулой тропонина С. После этого тропонин-тропомиозиновый комплекс подвергается конФормационным изменениям и сопровождается передвижением молекулы тропомиозина более глубоко в борозду между двумя молекулярными нитями актина. После этого открывается активный центр на молекуле актина, таким образом, позволяя головке миозиновой молекулы присоединиться к активному центру на молекуле актина”
60. в чем заключается роль ионов водорода в процессе сокращения мышечного волокна?
Существенным моментом сокращения является то, что в ре-зультате АТФ-азной реакции миозина происходит выделение ионов водорода,т.е. закисления среды. Рассчитано, что если только половина головок молекул миозина осуществляет одиночную АТФ-азную реакцию, рН понизится до 3,7. Полагают, что между ионами водорода и кальция происходит конкуренция за карбоксилаты в тропонине С.
Выделение ионов Н+ в результате АТФ-азной реакции миозина приводит к смещению равновесия между ионизированной и неионизированной формами карбоксилатов в сторону образования последней. В свою очередь это ведет к ослаблению взаимодействия ионов Са2+ с тропонином и к его диффузии в саркоплазму с последующим активным транспортом в саркоплазматический ретикулум.
Кроме того, процессы сокращения ведут к уменьшению концентрации АТФ и фосфора, что активирует систему ресинтеаа АТФ. В итоге концентрация ионов Са2+ уменьшается и повышается степень ионизации карйоксилатов тропонина С. Следователь¬о, при отсутствии ионов Са2+ тропонин-тропомиозиновый комплекс переходит в "выключенное" состояние, препятствующее акто-миозиновому взаимодействию. Следовательно, колебания концентрации Н+ могут оказывать регуляторное влияние на процесс сокращения мышцы.
61. в чем заключается механизм расслабления мышц?
В начале расслабление идёт пассивно за счёт эластических компонентов мышцы (сухожилий, связок), а затем активно. Главным моментом в активном расслаблении является секвестрация (sequestrare, лат. – отделять) кальция миоплазмы в его хранилища, т.е. в СПР. Удаление кальция из миоплазмы производит кальциевый насос, главной частью которого является Са++-активируемая Mg++-зависимая АТФ-аза, находящаяся в мембране элементов СПР. В активации этой АТФ-азы принимают участие:
1) большая концентрация кальция в присутствии ионов магния;
2) фосфаты, которые образуются при гидролизе АТФ;
3) белок кальсеквестрин, который находится в мембране цистерн СПР и принимает участие в секвестрации.
Са++-активируемая АТФ-аза расщепляет АТФ, высвобождается энергия и кальций активно нагнетается против градиента концентрации в цистерны СПР. Концентрация кальция в миоплазме становится равной примерно 10-7 М, головки миозина отсоединяются от активных участков актина и мышца расслабляется.
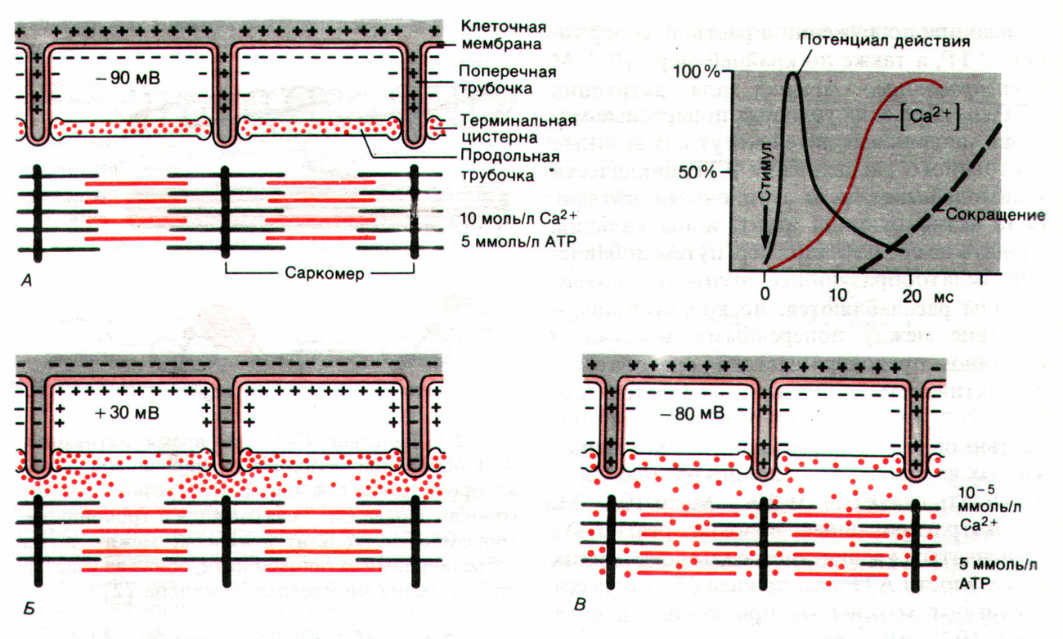
62. какие процессы лежат в основе энергообразования в мышце?
Основная энергетическая система, обеспечивающая работу – анаэробно-гликолитическая. В основе этого пути энергообеспечения лежит процесс гликолиза. Гликолиз – это сложный ферментативный процесс последовательных превращений углеводов (гликогена мышц и глюкозы) протекающий в саркоплазме мышечного волокна без потребления кислорода и сопровождающийся накоплением молочной кислоты. Следовательно, энергетические субстраты гликолиза, необходимые для образования АТФ – гликоген мышц и частично глюкоза, поступающая в мышцы с кровью. Конечный продукт гликолиза – молочная кислота, которая накапливается в мышцах с большой скоростью, поступает в кровь и вызывает выраженное снижение рН. Процесс гликолиза можно разделить на три стадии:
1. Подготовительная стадия. Происходит активация углеводов и образование субстрата биологического окисления.
2. Биологическое окисление и образование первичных макроэргических соединений.
3. Восстановление пирувата с образованием лактата.
Пусковыми ферментами гликолиза являются фосфорилаза и гексокиназа, расщепляющие соответственно гликоген и глюкозу. Активность этих ферментов зависит от содержания в саркоплазме АДФ и неорганического фосфора, ионов Са2, освобождающихся при мышечном сокращении, и концентрации катехоламинов в крови.
Первая стадия начинается с реакции фосфоролиза гликогена или с активации глюкозы при помощи АТФ с участием гексокиназы. В том и другом случае образуется глюкозо-6-фосфат, который превращается во фруктозо-6-фосфат. Это соединение активируется при помощи АТФ и фермента фосфофруктокиназы, и образуется фруктозо-1,6-дифосфат. Под действием фермента альдолазы это соединение распадается на 2 молекулы фосфоглицеринового альдегида.
На 2-й стадии протекает окисление 3-фосфоглицеринового альдегида с участием НАД-дегидрогеназы и фосфорной кислоты. При этом образуется 1,3-дифосфоглицерат – макроэргическое соединение. Далее происходит перефосфорилирование этого соединения с АДФ и образование АТФ путем субстратного фосфорилирования. В следующей реакции остаток фосфата из положения 3 переносится в положение 2, а затем происходит дегидратация 2-фосфоглицерата. Это приводит к образованию фосфоэнолпирувата с макроэргической связью, появление которой обусловлено электронной перестройкой молекулы. Затем вновь происходит реакция субстратного фосфорилирования – перенос макроэргического остатка с фосфоэнолпирувата на АДФ. Образуется еще одна молекула АТФ и пируват (пировиноградная кислота).
На заключительной 3 стадии гликолиза водород, отнятый НАД-дегидрогеназой от 3-фосфоглицеринового альдегида переносится от НАД∙Н2 на пируват, который при этом превращается в лактат (молочную кислоту) при участии фермента лактатдегидрогеназа. Кофермент НАД освобождается таким образом от протонов и электронов водорода и может участвовать в окислении новых молекул 3-фосфоглицеринового альдегида.
Биологическая роль гликолиза заключается в образовании промежуточных макроэргических соединений: дифосфоглицериновой и фосфоэнолпировиноградной кислот. Под действием ферментов эти соединения отдают свои высокоэнергетические фосфатные группировки на АДФ и образуется АТФ.
ФГК ~ Ф + АДФ фосфоглицераткиназа АТФ + ФГК
(дифосфоглицериновая кислота и фосфоглицериновая кислота)
ЭПВК ~ Ф + АДФ пируваткиназа АТФ + ПВК
(фосфоэнолпировиноградная кислота и пировиноградная кислота)
Наивысшей скорости гликолиз достигает уже на 30 секунде и обеспечивает поддержание максимальной мощности упражнения в интервале от 30 до 90 секунд. Однако довольно быстрое исчерпание запасов гликогена мышц и резкое повышение концентрации молочной кислоты, образующейся в результате гликолиза, приводит к снижению активности ключевых ферментов и внутриклеточного рН, что приводит к падению скорости гликолиза и подключению аэробных процессов (дыхания).
Мощность процесса менее 750 кал/кг/мин. Мощность зависит в основном от скорости процесса и регулируется активностью ферментов фосфорилазы и фосфофруктокиназы. Мощность достигает максимума с 20-30 секунды, держится 1-2 минуты и постепенно снижается при поступлении кислорода в мышцы, а также при снижении рН.
Емкость процесса около 2-3 минут. Емкость зависит от запасов гликогена в мышцах, от возможностей буферных систем и от устойчивости ферментов к накоплению лактата.
Эффективность низкая – 30-40%, т.к. углеводы расщепляются только до лактата и большое количество энергии расходуется в виде тепла.
Лимитирующим ферментом гликолиза является фосфофруктокиназа. Увеличение активности этого фермента в 5 раз увеличивает валовый поток гликолиза в 1000 раз. Также значительно влияют на процесс гликолиза фермент фосфорилаза, накопление лактата, емкость буферных систем и запасы гликогена в мышцах.
В спорте гликолитическая система энергообеспечения является основной при выполнении физических нагрузок продолжительностью от 30 секунд до 2,5 минут (в данной случае бег на 800м. – 2 минуты) с предельной для этой продолжительности интенсивностью и составляет основу скоростной выносливости. А также обеспечивает возможность ускорения по ходу дистанции и на финише.
63. опишите классификацию гладкомышечных клеток.
Мультиунитарная гладкая мышца.
состоит из отдельных гладкомышечных клеток, каждая из которых, находится независимо друг от друга;
имеет большую плотность иннерваации;
как и поперечнополосатые мышечные волокна, снаружи покрыты веществом, напоминающим базальную мембрану, в состав которого входят, изолирующие клетки друг от друга, коллагеновые и гликопротеиновые волокна;
каждая мышечная клетка может сокращаться отдельно и ее активность регулируется нервными импульсами;
входят в состав цилиарной мышцы, мышц радужки глаза, мышцы поднимающей волос.
Унитарная гладкая мышца (висцеральная).
представляет собой пласт или пучок, а сарколеммы отдельных миоцитов имеют множественные точки соприкосновения. Это позволяет возбуждению распространяться от одной клетки к другой
мембраны рядом расположенных клеток образуют множественные плотные контакты (gap junctions), через которые ионы имеют возможность свободно передвигаться из одной клетки в другу
потенциал действия, возникающий на мембране гладкомышечной клетки, и ионные потоки могут распространяться по мышечному волокну, обеспечивая возможность одновременного сокращения большого количества отдельных клеток. Данный тип взаимодействия известен как функциональный синцитий
подобный тип гладкой мышцы представлен в стенках большинства внутренних органов, включая кишечник, желчевыводящие протоки, мочеточник и большинство кровеносных сосудов
64. в чем состоят особенности сокращения гладкой мышцы и его механизма?
Гладкие мышцы, как и поперечно-полосатые,
содержат миозин, с поперечными мостиками, гидролизующий АТФ, и для того, чтобы вызвать сокращение, взаимодействует с актином.
В противоположность поперечно-полосатым мышцам,
тонкие филаменты гладких мышц содержат только актин и тропомиозин и не содержат тропонин;
регуляция сократительной активности в гладких мышцах происходит благодаря связыванию Са + с кальмодулином, активирующим киназу легкой цепи миозина, которая фосфорилирует регуляторную легкую цепь миозина. Это приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков.
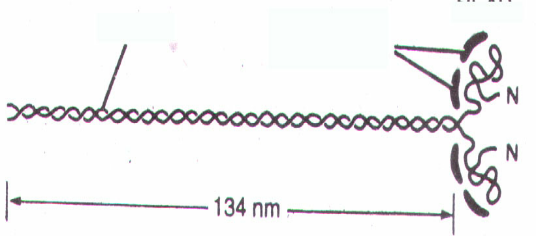 молекула
миозина
молекула
миозина
В гладкой мышце движение актомиозиновых мостиков является более медленным процессом
Распад молекул АТФ и высвобождение энергии, необходимой для обеспечения движения актомиозиновых мостиков происходит не так быстро как в поперечнополосатой мышечной ткани.
Экономичность энерготрат в гладкой мышце является чрезвычайно важным в общем потреблении организмом энергии, так как, кровеносные сосуды, тонкий кишечник, мочевой пузырь, желчный пузырь и другие внутренние органы постоянно находятся в тонусе.
Электромеханическое сопряжение осуществляется гораздо медленее, чем в поперечнополосатой мышце.
Сокращение типичной гладкой мышцы начинается спустя от 50 до 100 мсек после начала ее возбуждения, достигая своего максимума через 0,5 сек, а затем угасает в течение последую-щих 1-2 сек.
В гладкой мышце появление на сарколемме потенциала действия активирует фосфолипазу С и появлениие инозитол-3-фосфата, который связывается со специфическим для него рецептором, расположенным на кальциевом канале терминальной цистерны СПР. Это приводит к открытию этих каналов и выходу кальция из цистерны СПР.
Во время сокращения способна укорачиваться вплоть до 2/3 ее первоначальной длины (скелетная мышца от ¼ до 1/3 длины). Это позволяет полым органан выполнять свою функцию - изменять свой просвет от в значительных пределах.
Отношение актина к миозину в гладких мышцах (14-16: 1) намного больше, чем в скелетных мышцах (2:1). Тонкие филаменты гладких мышц прикреплены к плотным тельцам, а не к Z-линиям, и миозиновые поперечные мостики толстых филаментов могут взаимодействовать с тонкими филаментами на значительно большей протяженности их длины. Это обеспечивает гораздо более высокую степень укорачивания гладких мышц по сравнению с поперечно-полосатыми.
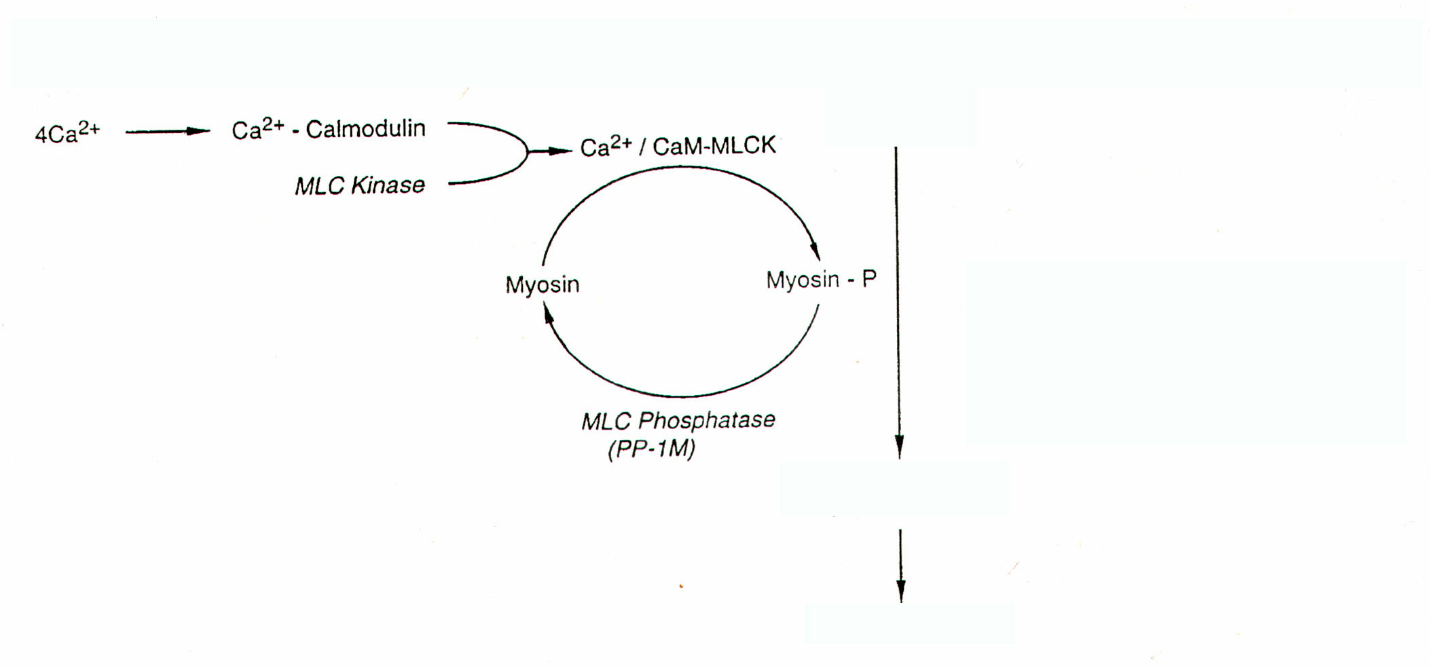
Актин- Активация миозиновой АТФазы – сокрашение
Миозиновые молекулы содержат 4 легкие цепи, две из которых связаны с головкой молекулы миозина. Головка миозиновой молекулы присоединяется к актину только после того как на ней фосфорилируются одна из легких цепей, получившая название регуляторной.
Фосфорилирование легкой цепи миозина катализируется киназой легких цепей миозина (КЛЦМ), которая активируется кальмодулином после его взаимодействия с ионами кальция.
Дефосфорилирование легких цепей миозина осуществляется фосфатазой легких цепей миозина (ФЛЦМ).
Скорость укорочения гладкого миоцита (то есть скорость циклинга акто-миозиновых мостиков) зависит от интенсивности фосфорилирования легких цепей миозина. При преобладании процесса дефосфорилирования над процессом фосфорилирования гладкая мышца расслабляется.
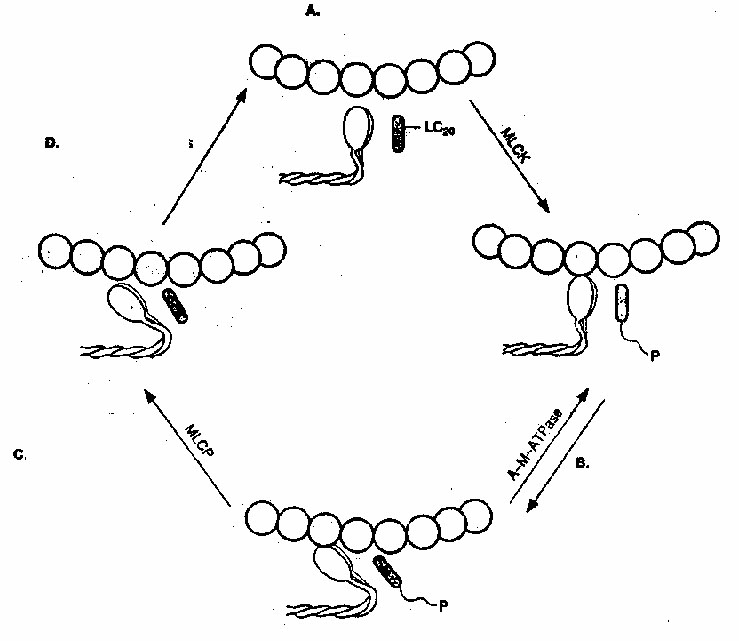
А - Расслабленная мышца
Б - Укорочениеи развитите силы
С – Поддержание силы «мостики на замке»
Д - Снижение уровня кальция
65. Особенности мембранного потенциала гладких мышц.
Потенциал покоя. Величина разности потенциалов на мембране гладкомышечных клеток существенно различается в зависимоти как от типа гладкой мышцы так и от тех условий, в которых она находится. Обычно в состоянии покоя мембранный потенциал гладкомышечной клетки нестабилен и находится в пределах от –30 до –50 мВ, что на 30 мВ меньше, чем в скелетной мышце. Потенциал действия унитарной мышцы. Потенциал действия в унитарной (висцеральной) гладкой мышце возникает так же как и в скелетной мышце. В висцеральных гладких мышцах потенциал действия различается по форме, амплитуде и продолжительности. Он бывает (1) в виде спайка или (2) потенциал действия, имеющий плато. Типичный spike-потенциал, характерен для гладкой и скелетной мышцы. Его продолжительность от 10 до 50 мсек. Данный потенциал возникает при нанесении на гладкую мышцу электрического, химического раздражения, а также растяжения. Кроме того, потенциала действия подобного типа может возникать спонтанно. Потенциал действия, имеющий плато, своим началом напоминает spike-потенциал. Однако сразу после быстрой деполяризации начинается быстрая реполяризации. Однако она задерживается вплоть до 1000 мсек. Так формируется плато потенциала действия. Во время плато гладкая мышца длительное время остается укороченной. Подобный тип возбуждения имеет место в гладкой мышце мочевого пузыря, матки и др. Необходимо отметить, что в мембране гладкомышечной клетки обнаружено гораздо большее количество потенциалзависимых кальциевых каналов, чем в мембране поперечнополосатых мышечных волокон. Более того, ионы натрия играют малую роль в гененерации потенциала действия. Вместо них большое значение в генерации потенциала действия принадлежит потоку ионов кальция внутрь гладкомышечной клетки. Однако кальциевые каналы открываются значительно медленее, чем натриевые каналы, но остаются открытыми значительно дольше. На основании этого можно понять почему потенциал действия гладкой мышцы развивается в течение столь длительного времени. Другой важной задачей входящего во время потенциала действия кальция является их прямое влияние на сократительный аппарат клетки. Некоторые гладкомышечные клетки обладают способностью к самовозбуждению, то есть способны генерировать потенциал действия без воздействия внешнего раздражителя. Это часто связано с периодическими колебаниями мембранного потенциала. Очень часто подобная активность наблюдается в гладкой мышце кишечника. Медленные волновые колебания мембранного потенциала не являются потенциалом действия. Одним из возможных механизмов, объясняющих появление этих волновых колебаний мембранного потенциала, является периодическая активация и затухание активности натрий-калиевого насоса. Разность потенциалов на мембране гладкомышечной клетки увеличивается во время активации Na/K насоса и уменьшается при ее снижении. Другой возможной причиной данного явления является ритмическое увеличение или снижение проводимости ионных каналов. Физиологическое значение медленных колебаний мембранного потенциала состоит в том, что они могут инициировать появление потенциала действия. Это возникает в том момент когда во время медленной волны разность потенциалов на мембране клетки снижается до –35 мВ. При этом, как правило, успевает возникнуть несколько потенциалов действия. Следовательно, медленные волны можно назвать пейсмекерными волнами и, таким образом, становится понятным каким образом они обуславливают ритмические сокращения кишки. Одним из важных раздражителей, инициирующих сокращение гладких мышц, является их растяжение. Достаточное растяжение гладкой мышцы обычно сопровождается появлением потенциалов действия. Таким образом, появлению потенциалов действия при растяжении гладкой мышцы способствует два фактора: (1) медленные волновые колебания мембранного потенциала, на которые наслаивается (2) деполяризация, вызываемая растяжением гладкой мышцы. Данное свойство гладкой мышцы позволяет ей автоматически сокращаться при растяжении. Например, во время переполнения тонкого кишечника возникает перистальттическая волна, которая и продвигает содержимое.
66. Назовите пути ресинтеза АТФ в клетке.
Хорошо известно,что сокращение мышцы зависит от энергии АТФ, и для постоянного сокращения мышцы необходим ее постоянный ресинтез. Для этого в организме есть ряд путей:
1. Самый быстрый путь это креатинкиназная реакция, названная так из-за фермента креатинкиназы, при участии которого она осуществляется. В мышцах содержится креатинфосфат (КФ), который может передавать свою фосфатную группу со всем запасом энергии связи с креатином на АДФ.
К Ф+ АДФ= К+АТФ
(это типичная фосфотрансферазная реакция).
Как только уровень АТФ начинает снижаться, сразу же запускается эта реакция, что обеспечивает ресинтез АТФ. Поэтому естественно, что при мышечной деятельности содержание КФ снижается в большей степени, чем уровень АТФ.
2. Следующим путем ресинтеза АТФ является гликолиз, то есть происходящее без участия кислорода (анаэробно) окисление глюкозы до молочной кислоты.
3. Важным источником АТФ является окислительное фосфорилирование. Таким образом образуется 95% энергии потребляемой мышцей в процессе сокращения. Именно этот путь способен обеспечить энергией длительное мышечное сокращение. Субстратами для цикла Кребса могут быть углеводы, жиры и белки.
67. Теплообразование в мышце и его деление на фазы.
В процессе мышечного сокращения часть используемой энергии превращается в механическую работу, а часть рассеива-
ется в виде тепла. Следовательно, по количеству выделенного
тепла можно судить об энергетических затратах, используемых
на мышечное сокращение.
Используя микротермопару Хилл измерил теплообразование в акте одиночного мышечного сокращения. Он подразделил его на 2 основные фазы:
1. Начальное теплообразование,
2. Запаздывающее, или восстановительное теплообразование.
Начальное теплообразование короче запаздывающего в 1000 раз и делится на : а) тепло активации, б) тепло укорочения,
в) тепло расслабления.
Тепло активации - включает в себя то тепло, которое выделяется сразу после нанесения раздражения. В первую очередь
- это тепло, которое высвобождается при активации Na-K насоса.
Тепло укорочения - обусловлено самим сократительным процессом (активация миозина, актомиозиновое взаимодействие, отсоединение актина от миозина).
Тепло расслабления - это тепло, которое высвобождается во время работы Са2+ насоса СПР, мембраны мышечного волокна и натрий-калиевого насоса.
Фаза начального теплообразования обеспечивается за счет анаэробных гликолитических процессов и за счет запасов АТФ.
Запаздывающее, или восстановительное теплообразование длится несколько минут после расслабления. В это время восстанавливается резерв АТФ: на 90% за счет окислительного фосфорилирования и на 10% за счет гликолиза.
68. Что такое синапс? Классификация синапсов.
Синапс - это структура, которая обеспечивает морфофункциональную связь окончания аксона или какой-то другой части клетки с дендритом, телом или аксоном другого нейрона или в некоторых случаях с мышечной или железистой клеткой.
С точки зрения функции синапсы можно разделить на химические и электрические.
Электрические синапсы - это образования, в которых возбуждение передается в форме "скачка" с одной клетки (окончания пресинаптической клетки) на мембрану другой клетки (постсинаптическую). Мембраны этих клеток плотно соприкасаются друг с другом. Поэтому между ними существует низкое сопротивление, благодаря которому ионы проходят относительно легко. Электрические синапсы встречаются, например, между некоторыми нейронами в латеральных вестибулярных ядрах продолговатого мозга.
Однако, большинство синапсов - это химические синапсы, т.е. передача сигнала в них происходит химическим путем. Нейромышечный синапс - это специализированная область, где окончание двигательного нейрона оканчивается на мембране поперечно-полосатого мышечного волокна.
Контакт между окончанием аксона нейрона вегетативной нервной системы и гладкой мышцей или сердечной мышцей является менее специализированным образованием по сравнению с нервно-мышечным синапсом, и передача в нем в большей степени является диффузным процессом.


 Классификация
химических синапсовпо месту своего
образования
Классификация
химических синапсовпо месту своего
образования
Нервно-мышечные Нервно-железистые Нервно-нервные