

2.2. Гликопротеины и протеогликаны
Углевод-белковые комплексы данных сложных белков отличаются друг от друга по ряду признаков представленных в таблице 5:
Таблица 5
Отличия гликопротеинов и протеогликанов
Гликопротеины |
Протеогликаны |
Характер простетической группы |
|
Представлены: моносахаридами и олигосахаридами. Олигосахариды – это небольшие гетерополисахариды нерегулярного строения. Могут иметь разветвленную структуру, включать от 2-х до 10 (иногда более) мономеров Мономеры олигосахаридов представ- лены: глюкозой, галактозой, маннозой, фукозой, N-ацетилгалактозами- ном, N-ацетилглюкозамином, сиаловой кислотой (N-ацетилнейраминовая кислота) |
Представлены: гликозаминогликанами (старое название мукополисахариды). Это высокомолекулярные линейные гетерополисахариды,. Молекулярная масса колеблется от 105 до 4·106 Построены из повторяющихся дисахаридных единиц, которые чаще состоят из гексуроновой кислоты и галактозамина или N-ацетилпроизводного глюкозамина Реже встречаются фукоза, манноза и сиаловые кислоты |
Характер связи в углевод-белковом комплексе |
|
Связь прочная, ковалентная. Олигосахаридные цепи прочно (ковалентно) присоединены к полипептидной основе |
Гликозаминогликаны обычно ковалентно связаны с белками, но возможна и слабая (адсорбционная) связь |
Характер образования ковалентной связи в обеих группах имеет общие черты. Углеводная часть может быть связана с белком за счет гидроксильных групп серина или треонина – это «О»-гликозидная связь или за счет амидной группы аспарагина – это «N»-гликозидная связь |
|
Характер среды |
|
Нейтральный или слабокислый за счет присутствия сиаловых кислот |
Выраженный кислый характер за счет большого количества уроновых кислот и серной кислоты, имеют значительный отрицательный заряд |
Место нахождения |
|
Во всех органах, тканях и биологических жидкостях |
В основном находятся в межклеточном веществе соединительной ткани |
Белок составляет от массы молекулы |
|
80–90 % |
5–10 % |
Все моносахариды, входящие в состав протеогликанов и гликопротеинов синтезируются в организме из глюкозы.
2.2.1. Общие черты синтеза углеводной части гликопротеинов и протеогликанов
Синтез гетерополисахаридов происходит при участии гликозилтрансфераз. Донорами моносахаридных остатков служат нуклеотид сахара (например, уридиндифосфат~глюкоза – УДФ~глюкоза). «Активирование» моносахаридов осуществляется в цитозоле при участии соответствующих нуклеозидтрифосфатов. Образование УДФ ~ галактозы требует двух реакций:

 ..
..
Реакции гликозилирования протекают в просвете элементов комплекса Гольджи. Нуклеотидсахара (УДФ~галактоза, ГДФ~манноза, ЦМФ~нейраминовая кислота) поступают в цистерны аппарата Гольджи при участии систем-переносчиков по механизму антитранспорта в обмен на соответствующий нуклеотид: УМФ, ГМФ, ЦМФ. Например, УМФ образуется из УДФ~галактозы следующим образом:
Галактозил
трансфераза
Нуклеозидфосфатфосфатаза

Гликозилтрансферазы – это очень большая группа ферментов, с помощью которых образуются разнообразные гетерополисахариды. Каждый фермент строго специфичен и к донору моносахарида (например, УДФ~глюкоза, ГДФ~манноза, ЦМФ~нейраминовая кислота и др.), и к его акцептору (белку или растущей олигосахаридной цепи гликопротеина).
Не смотря на то, что синтез гетерополисахаридов не относится к матричным, результат получается сходным в том отношении, что образуются полимеры олигосахаридов не с произвольной, а со строго определенной последовательностью мономеров (моносахаридов и их производных – аминосахаров и ацетилированных гексозаминов).
Синтез углеводной части гликопротеинов, соединенной с белком «О»-гликозидной связью отличается от механизма синтеза олигосахаридов и гетерополисахаридов, присоединенных «N»-гликозидной связью.
При синтезе гликопротеинов и протеогликанов с «О»-гликозидной связью, ОН–группа серина или треонина белковой молекулы является основой, на которой постепенно наращиваются цепи гетерополисахаридов.
В случае синтеза гликопротеинов с «N»-гликозидной связью (основой является амидная группа аспарагина), участвует промежуточный носитель олигосахарида – долихолфосфат – вещество изопреноидной природы (Dol–p):
Остаток
фосфорной кислоты
Долихолфосфат
(n
= 17–20 единиц изопрена)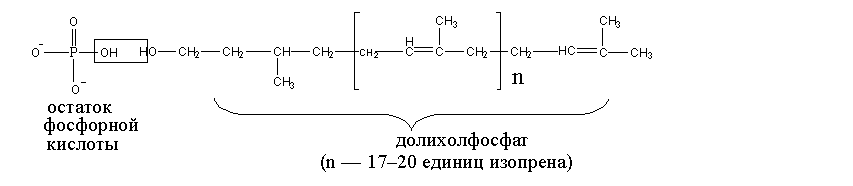
Гидрофобный конец долихолфосфата погружен в липидный слой мембраны аппарата Гольджи или эндоплазматического ретикулума (ЭПР), а на выступающем гидрофильном конце (остатке H3PO4) наращивается олигосахарид при действии специфических гликозилтрансфераз.
Образовавшийся олигосахарид из десятка и более моносахаридов целиком переносится с долихолфосфата на амидную группу аспарагина в составе полипептидной цепи белка. Затем происходит доработка (модификация) олигосахаридов. Реакция переноса катализируется мембраносвязанным ферментом олигосахарид-трансферазой. Данный фермент узнает и переносит любой углеводный фрагмент с общей структурой (R-(глюкоза-N-ацетил)-2-Р-Р-Dol. Затем гликозилирование белка происходит по остатку аспарагина трипептидной последовательности Асн–Х–Сер/Тир, где «Х» – любая аминокислота за исключением, вероятно, аспарагиновой кислоты и пролина (в структуре гликопротеинов). При этом предпочтительно используется трипептид, содержащийся в β-цепи (кросс β-форма). Лишь треть остатков аспарагина в белке реально подвергаются гликозилированию.
Долихолдифосфат (Dol–p-p) подвергается дефосфорилированию под действием фосфатазы. Долихол-Р может вновь служить акцептором для синтеза следующей молекулы олигосахарид–р-р–долихол.
Процессинг (или модификация) олигосахаридной цепи, связанной с белком включает ранние стадии, которые сводятся к удалению концевых одного или двух остатков глюкозы и, возможно, одного или четырех остатков маннозы при участии ферментов глюкозидазы-I, глюкозидазы-II, и различных маннозидаз. Очень важным завершением ранней стадии является реакция присоединения глюкозо-N-ацетил-1-фосфата к 6-му углеродному атому одного или нескольких специфических остатков маннозы. Реакция катализируется ферментом глюкозо-N-ацетил-фосфотрансферазой, использующей в качестве донора УДФ~глюкозо-N-ацетил. Образуются продукты: УМФ и глюкозо-N-ацетил-РО4–-6-манноза-белок. Этот новый концевой фрагмент олигосахарида в следующей реакции (фермент фосфодиэстераза) освобождается от глюкозо-N-ацетил, а остатки маннозы в 6-ом положении остаются фосфорилированными.
Рецепторы для маннозо-6-фосфата, локализованные в аппарате Гольджи, связывают маннозо-6-фосфат этих белков. Одна из функций маннозо-6-фосфата в аппарате Гольджи – распознавать и направлять ферменты лизосом к этим органеллам. При снижении активности глюкозо-N-ацетилфосфотрансферазы или отсутствии (молекулярная болезнь) в клетках накапливается множество различных видов недеградированных молекул гликопротеинов. В то же время в пробах сыворотки больных этой болезнью можно обнаружить высокую активность лизосомных ферментов. Это свидетельствует о том, что синтез ферментов происходит, но они не достигают предназначенных для них внутриклеточных органелл.
Поздняя стадия процессинга олигосахаридной цепи заключается в присоединении ко второй олигосахаридной цепи разветвленной структуры (биоантенной) второго глюкозо-N-ацетильного остатка (фермент глюкозо-N-ацетилтрансфераза II) и остатков фукозы, галактозы и сиаловой кислоты (N-ацетилнейраминовой) в различных положениях при участии ферментов фукозил-, галактозил- и сиалилтрансферазами.
Субклеточная локализация. Главными структурными компонентами клетки, участвующими в процессах синтеза белковой части и гликолизировании являются эндоплазматический ретикулум и аппарат Гольджи. Присоединение олигосахарида протекает в шероховатом эндоплазматическом ретикулуме во время и после синтеза белка. Удаление глюкозы и некоторых из периферийных остатков маннозы также происходит в ЭПР и затем переносятся в цистерну Гольджи для последующих реакций гликозилирования.