

Индукцияланған мутанттардан бөлініп алынған антибиотиктер продуценттері штаммаларының
ӨНДІЛІП
Антибиотиктің аттары |
Мутацияны туғызған фактор |
Штаммалардың енімділігі (актиатіліКтің бірлігі) |
|
|
|
баста пқылары |
жаңалары |
Пенициллин Стрептомицин Хлортетрациклин |
Р, УФ, АИ, ЭИ Р, УФ
Р, УФ, ЭИ |
100 250
600 |
5200 4200
3500 |
Белгілер: Р — рентген сәулесі, УФ — ультракүлгін сәуле, ЭИ — эти-ленимин, АИ — азотты иприт. |
|||
мен селекция арқылы алынған штаммалардың активтілігі жөніндегі деректер келтірілген.
Селекцияда мутагеннің бір рет эсер етуімен шектелмейді. Сүрыптап алынған үздік штамма мутагенді қайталап қолдан-ғаннан кейін қайталап сүрыптаудың объектісіне айналды, солай жалғастырыла береді. Осындай көп рет сұрыптау селекцияның әрбір сатысында өнімділікті арттырады, сондықтан ол көп са-тылы сүрыптау деп аталды. Көп сатылы сұрыптау мысалы 101-суретте келтіріліп отыр.
Индукцияланған мутацияларды насекомдар селекциясында пайдалану. Өсімталдығы темен жануарлар селекциясында ин-дукци-яланған мутациялар алу әзірше өте шектеулі болып отыр, бірақ насекомдарда (жібек көбелектері, бал арасы) мүның практикалық маңызы болады.
Мысалы, түт жібек көбелегінде еркек жынысты пілләлардан алынатын жібек ұрғашы жынысты пілләларға қарағанда 20—ЗО°/о көп болады.
Таяуда В. А. Струнников жібек көбелегінің үрғашысыз түқымын шығарды, оның жүлдыз қүрттарының бәрі еркек
ЮІ-сурет.
Сатылы селекция
әдісі
бо
йынша
штаммаларды
—
хлортетрацик-
линнің
продуценттерін алу. Штам-
малардыи,
нөмірі керсетілген жоне
жақшада
активтілік бірлігі бойынша
(мкг/мл)
өнімділігі берілген.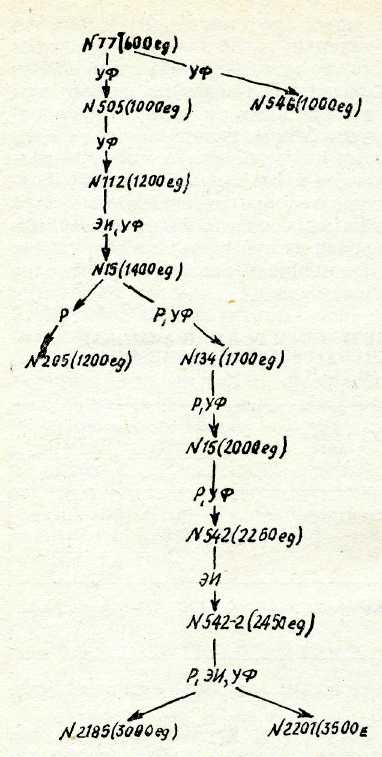
Асыл түқымды аталық-тың екі Х-хромосомаларының әрқайсысының мынадай // мутациясы —-~т болады, мұнда мен /2 өзара аллельді емес, демек бүл аталық өміршең болады. Ол U неме-се k бойынша гетерозиготалы ұрпағында өміршең ер-ек және гемизиготалы (эмбрион кезінде өліп қалатын) ұрғашы береді. Сонымен мүұндай аталықтың есіп шығатын барлық ұрпағы—гусе-ницалар еркек жынысты болады. Жібек көбелегінІЕ «ұрғашысыз» түқымының шығарылуы селекцияда генетикалық анализ принцип-тері мен әдістерін пайдала-нудың тамаша мысалы.
3. Полиплоидия
Полиплоидия есімдіктер селекциңсы үшін өзгергіштіктің аса бағалы қайнар кезі болып табылады, ол мә-дени өсімдіктер селекциясында ете маңызды роль атқарады.
Селекцияда автополип-лоидияны пайдалану. Соңғы жылдарда теориялық зерттеулердің табыстары кептеген перспективалы полиплоид формаларын алуға мүмкіндік жасады. Колхицинды пайдалану полиплоид-тарды алуды жеңілдетті. Диплоидтан тетраплоидқа хромосомалардың санын екі есе үлғайту көбінесе клеткалардың көлемімен бөліну қарқынының өзғеруімен бірге жүреді, бүл есімдіктің көлемінің және оның органдарының, түқымның мөлшері мен массасының, олардың химиялық қүрамының және т. б, өзгеруіне әкеп соғады. Мәселен, В. С. Федоров селекциялаған тетра плоидты қара бидайдың Ленинград Тетрасы сортының 1000 дәнінің массасы 55—56 г, ал осы. сорттың -диплоиды — 29 г болды.
Полиплоидтандыру кейбір жағдайларда бағалы химиялық заттардың көп болуын (мәселен, фармацевтика өнеркәсібі үшін маңызды көкнәр апиынындағы, кейбір органикалық қосылыстар, жүгерідегі Л витамині) қамтамасыз етеді немесе, керісінше, кей-бір адам үшін қажетсіз қандай да болмасын қосылыстардың синтезделуін азайтады (мысалы, полиплоидты қант қызылша-сындағы азот қосылыстары). Полиплоидтар бүлардан басқа да бағалы белгілерге ие болады, мысалы, ауруларға қарсы төзімді-ліктің жоғарылауы т. б.
Сонымен бірге қолдан алынатын автополиплоидтардың фертилділігі темен болады. Полиплоидта бастапқы формаға қарағанда әрбір түқым ірі, бірақ өсімдІктегі түқым саны, әдет-те, аз болады. Мүның себебі негізінен мейоздың бұзылуына байланысты (13-тарауды қараңыз). Алайда бүл кедергілерді жеңуге болады. Полиплоид дайын сорт емес, ол одан әрі мұқи-ят селекцияны қажет етеді.
Казіргі уақытта шаруашылыққа қажетті бағалы полиплоидтар бірқатар ауыл шаруашылық дақылдарынан: қант қызыл-шасы, тары, көкнәр, зығыр, шалқан, жүгері, сәндік өсімдіктер т. б. алынды. Ауыл шаруашылық өндірісінде триплоидты формаларды пайдалану үлкен тиімділік берді. Триплоидты өсімдік-тер, әдетте, стерильді болады. Бірақ олар, әдетте, күшті дамуы арқылы ерекшеленеді және сондықтан вегетативтік массасы жоғары өнімді болады. Атап айтқанда, мысалы тамыр жемістерінің өте ірі болуы арқасында триплоидты қант қызылшасы ауданның бір өлшеміне шаққанда диплоидқа қарағанда 10— 20% артық қант өнімін береді, бірқатар елдерде ол егіс келе-мінің 60—100%-тін алады. Триплоидты өсімдіктерді қызылша-ның диплоидты және тетраплоидты формаларын шағылыстыру жолымен алады. СССР-де түқымсыз триплоидты қиярлардың перспективалы жоғары енімді формалары алынды.
Кейбір автополиплоидты формаларда қажетті ерекшеліктер-мен қатар қажетсіз ерекшеліктер де болатынын ескеру керек. Мысалы, кейде клеткалардағы судың мөлшері көп болатыны байқалады, б9л қуаңшылыққа және аязға төзімділіктің азаюы-на соқтырады. Сондықтан полиплоидты формаларды жасау қатаң сүрыптаумен бірге жүруі тиіс.
Аллополиплоидтарды селекцияда пайдалану. Аллополипло-идтарды селекцияда пайдалану полиплоидты формалардың артықшылығымен комбинативті өзгергіштікті үйлестіруге мүм-кіндік береді. Мәселен, В. Е. Писарев бидай (2я = 42) мен қара бидайды (2/г=14) шағылыстыру жолымен амфидиплоид (2п = = 56) жасады, оған Triticale (бидай—Triticurn, қара бидай Seca-. le) деген ат берді. Triticale есімдігі қара бидай сияқты қара топырақты емес, алқап жағдайында жақсы жетіледі, белок қорын көп жинайды, бидай сияқты, тат ауруы мен ақ ұнтақ ауруына төзімді келеді.
Анеуплоидтарды селекцияда пайдалану. Анеуплондты орга-низмдердің әдетте, тіршілік қабілеті мен фертильділігі төмен болады, сондықтан өсімдік шаруашылығында практика жүзінде қолданылмайды. Бірақ оларды қазір өсімдіктер селекциясында бір генотиптің (түрдің) хромосомаларын басқа генотиптің (түр-дің) хромосомаларымен алмастыру әдісін пайдалануға барған сайын кеңінен қолдануда. Осы әдістің көмегімен донордың хро-мосомасымен бірге реципиентке қалаған гендердіц жиынтығы беріледі. Бүл нулли немесе моносомиктерді пайдалану арқасында мүмкін болады (13-тарауды қараңыз). Мәселен, бидай-дың Чайниз Спринг сортында әрбір хромосома бойынша мо-носомиктер алынды. Осындай моносомиктерді басқа сорттармен шағылыстыра отырып, бүл сорттың жекелеген хромосомаларын донор сорттың хромосомаларымен алмастырды. Мүндай шағылыстырулардың Ғгде донор мен реципиент хромосомалары бойынша гетерозиготалар алынады, ал одан әрі донор хромосомаларыныц бір жүбынан басқа бүкіл хромосомалары Чайниз Спринг реципиентіне ие болатын генотипті бөлуге болады. Жекелеген хромосомаларды алмастыру енімділікті, сабағының биік-тігін, жатып қалуға төзімділігін және т. б. езгертеді. Осы әдіспен Nicotiana таЬасшп түрінің геномында темекі моза-икасының вирусына сезімтал хромосомалардың біреуін осы ауруға төзім-ді генге ие N glutinos хромосомасына алмастырудың мүмкіндігі болды.A * AСонымен, селекцияда түқым қуалайтын өзгергіштіктіц екі типі де — комбинативті және мутация пайдаланылады. Осы немесе басқа типтің өзгергіштігін сүрыптау үшін пайдалану объектініц биологиясымен және селекционер алдында түрған мақсаттармен анықталады.
32-тарау. ШАҒЫЛЫСТЫРУ ЖҮЙЕЛЕРІ
Тұқым қуалайтын өзгергіштіктіц болуы әр түрлі шағылыстыру жүйелерініц жолымен бір организмде нақты тұқым қуалайтын белгілерді үйлестіруге, сондай-ақ үнамсыз қасиеттерден құтылуға мүмкіндік береді. Селекцияда комбинативтік өзгер-гіштікті пайдаланудыц қажетті шарты шағылыстыру үшін фор-маларды сүрыптау болып табылады.