

7.4.3. Фотопериодизм 7.4.3,1. История открытия фотопериодизма
В двадцатые годы XX в. американские исследователи У. У. Гарнер и Г. А. Аллард изучали влияние длины светового дня на табак. Объектом их исследований был табак сорта «Maryland Mammoth». Этот сорт отличался более крупными размерами и поздним цветением, поэтому для сбора семян приходилось доращивать растения в теплице. Гарнер и Аллард выяснили, что этот сорт цветет только при коротком дне, экспериментальное укорочение длины дня также вызывало цветение, т.е. табак «Maryland Mammoth» — типичное растение короткого дня (большинство сортов табака — нейтральные растения). Гарнер и Аллард пробовали прерывать светлое время суток периодом темноты, а ночное время — вспышками света. Прерывание дня не дало результата, прерывание ночи задерживало цветение, и чем ближе к середине ночи проводили освещение, тем эффективнее была обработка. Для предотвращения цветения достаточно нескольких минут ночного освещения (рис. 7.34).
Изученный феномен получил название эффекта прерывания ночи. Был сделан вывод, что для оценки длительности дня важно не светлое (дневное) время, а продолжительность ночи. Условно каждое короткодневное растение можно было назвать «растением длинной ночи», а каждое длиннодневное — «растением короткой ночи». (Впрочем, такой вывод оказался преждевременным.)
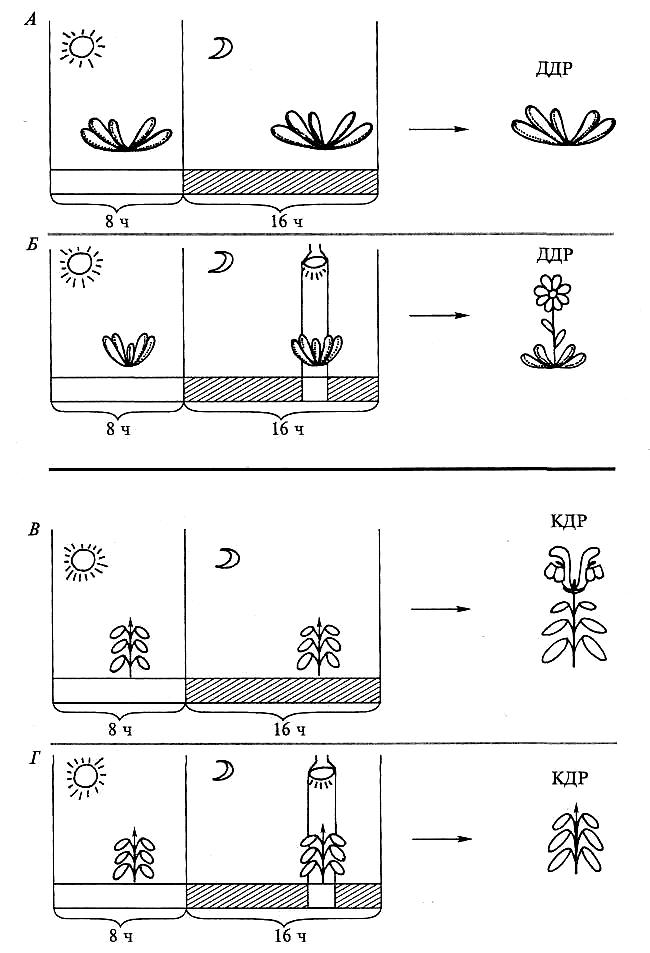
Рис. 7.34. Эффект прерывания ночи:
А — контроль: длиннодневное растение в условиях 8-часового дня/16-часовой ночи вегетирует; Б — опыт: 16-часовая ночь прервана 5 минутами освещения; длиннодневное растение цветет; В — контроль: короткодневное растение в условиях 8-часового дня/16-часовой ночи цветет; Г — опыт: 16-часовая ночь прервана 5 минутами освещения; короткодневное растение вегетирует. На нижней полосе темное время суток показано штриховкой, периоды освещения — белым
7.4.3.2. Опыты м.Х. Чайлахяна
В 1935 г. в Институте физиологии растений АН СССР начались исследования М.Х.Чайлахяна. Для большинства экспериментов он использовал простейшее оборудование: темную плотную ткань и скальпель. Для решения вопроса об органе, воспринимающем длину дня, были взяты хризантемы (КДР). В летнее время, чтобы вызвать цветение, достаточно было в течение 10—15 дней в строго определенное время накрывать и открывать растения черной тканью. Если ткань снимали на 10 ч и менее, хризантемы цвели (положительный контроль). Если растения оставались на естественном 16-часовом дне, то они вегетировали (отрицательный контроль).
На свету оказывались листья и апикальные меристемы побегов, поэтому нужно было выяснить, какой из этих двух органов воспринимает световой сигнал. Чайлахян оставил на каждом растении по одному крупному листу и по одной точке роста. Чтобы избежать влияния листьев, образуемых меристемой, каждые 2 дня молодые листья удаляли (рис. 7.35).

Рис. 7.35. Опыт М.Х.Чайлахяна с короткодневным растением Chrysanthemum indicum (Сложноцветные), показывающий роль листьев в восприятии длины дня:
А — хризантема в условиях 16-часового дня вегетирует (отрицательный контроль); Б — хризантема при 8-часовом дне цветет (положительный контроль); В — меристема получает 8 часов света, лист — 16 часов света ежедневно: хризантема вегетирует; Г — меристема получает 16 часов света, лист — 8 часов света ежедневно: хризантема цветет
Накрывая черным футляром меристему, Чайлахян добивался того, чтобы листья оказывались на длинном дне (16 ч), а меристема получала короткий день. В другом варианте футляром закрывали лист: меристема оставалась на длинном дне, а лист освещался 8 ч. В результате меристемы образовали соцветия только там, где для листьев создавался короткий день. Ученый сделал вывод, что длина дня воспринимается листьями, далее преобразуется в сигнал и передается в меристему побега, где вызывает переход к состоянию цветения. Опыты с другими группами растений (ДДР, ДКДР, КДДР) подтвердили этот вывод.
Один из самых изящных экспериментов был проведен с периллой (Perilla nankensis) (очень живучее КДР из сем. губоцветные (Labiatae)). M.X.Чайлахян вырастил периллу на длинном дне (неиндуктивные условия), затем отрезал листья и поместил их во влажной камере на короткий день (индуктивные условия), а затем привил эти листья на нецветущие растения. Несмотря на низкий процент приживаемости листьев, в удачных вариантах листья стимулировали цветение периллы! Это означало, что даже отрезанные от растения листья могут воспринимать длину дня, вырабатывать и накапливать сигнальные вещества, вызывающие цветение. Оставляя почки в нижней части растения, а листья в верхней, М.Х. Чайлахян выяснил, что сигнал может распространяться сверху вниз, т. е. нельзя говорить о полярном распространении флорального стимула. Более того, периллу удалось аккуратно разрезать почти до кончика корня. Листья одной половины растения, получавшие день нужной длины, передавали сигнал цветения к почкам в другой половине. Этим опытом было доказано, что сигнал передается не только по стеблям, но и через корневую систему. Была измерена скорость распространения сигнала, и выяснилось, что она зависела от температуры: при 5 °С скорость сигнала падала.
Опыты с надламыванием ксилемных пучков и удалением наружного слоя флоэмы со стебля показали, что сигнал от листьев к меристеме передается по флоэме, т. е. по живым клеткам.
Чтобы растение восприняло длину дня как стимул к цветению, оно должно достигнуть определенного возраста. М.Х.Чайлахян называл этот возраст цветочно-спелым состоянием, т. е. внутренние факторы развития должны привести растение к определенному состоянию. После получения сигнала меристема не сразу образует видимые органы цветка. Этому предшествует период «скрытой подготовки» к цветению (эвокация по Чайлахяну).