

6.2.4. Поглощение ионов интактным растением стационарного состояния
Успех изучения поступления ионов на модельных системах (клетки водорослей, протопласты, везикулы, обессоленные отсеченные корни) не вызывает сомнений: выявлены транспортеры, включенные в перенос разных ионов, выяснены молекулярные, электрофизиологические и кинетические характеристики многих переносчиков и каналов.
Однако исследования подобного рода не дают возможности получить ответы на ряд важных вопросов:
• Как происходит поступление ионов в естественной среде, где действуют многие факторы, некоторые из которых могут достаточно быстро меняться (доступность воды и ионов, температура и другие факторы)?
• Каким образом взаимодействуют надземные и подземные органы, обеспечивая поглощение ионов, и как регулируется процесс поступления элементов минерального питания в целостном организме?
• Какова взаимосвязь между доступностью элементов минерального питания, их накоплением в тканях, ростом организма и реализацией программы полного цикла развития?
Исследования на интактных растениях, которые находятся в стационарном состоянии, т. е. имеют определенный статус минеральных элементов (содержание или концентрация отдельных элементов и их соотношение) и приспособлены к тем или иным условиям среды, позволяют частично понять некоторые особенности функционирования систем поступления в целом организме.
6.2.4.1. Кинетика поглощения ионов интактным растением
Для интактных растений, которые находятся в стационарном состоянии, изотерма поглощения имеет вид типичной кривой зависимости поступления от концентрации иона в растворе (рис. 6.30) и аналогична изотермам, полученным на модельных системах (гигантские клетки харовых водорослей или отрезанные, обессоленные корни), где отсутствует взаимодействие корень-побег (транслокация ионов из корней и поступление ассимилятов).
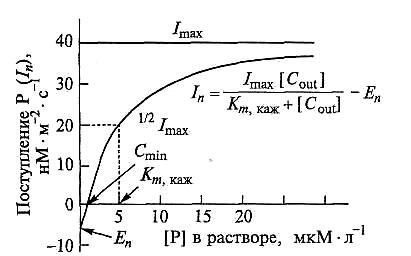
Рис. 6.30. Поток фосфора Iп в корни 18-дневных растений кукурузы в зависимости от его концентрации в растворе ([Р]) (по Барберу, 1988):
Imax – максимальный нетто-поток; Еn — нетто-выход при низкой концентрации иона в растворе; Cmin — концентрация, при которой потоки иона в корни и из корней равны; [Cout] — концентрация иона в растворе; Кт — константа Михаэлиса
В экспериментах, более приближенных к естественным условиям, для описания изотермы поглощения часто используют иные параметры. Это связано с тем, что поступление минеральных элементов в необессоленные корни, из которых ионы перемещаются в побеги, представляет собой чистый (нетто) поток In (расчет на поглощающую поверхность или на корни одного растения). Нетто-поток складывается из двух потоков: входа I (от influx) в корни и выхода Е (от efflux) из них. Именно величину In измеряют, когда поглощение оценивают по изменению концентрации наружного раствора. В связи с этим показатель Vmax в уравнении кинетики поглощения (6.3) заменяют на Imах — максимальный нетто-поток ионов в корни растений. Поэтому, когда поступление оценивается как поток ионов, In может быть определен по модифицированному уравнению Михаэлиса — Ментен (рис. 6.30). Imах отражает число мест переноса, включая рабочую поглощающую поверхность корня. Величина Кт,каж показывает, какие системы переноса (высоко- или низкоафинные) включены в транспорт иона; Еп и Cmin дают представление о соотношении входа и выхода. Следует подчеркнуть, что соотношение вход/выход при более высоких концентрациях может быть иным. Однако для характеристики поглощения интактными растениями из растворов питательных смесей в экспериментальных условиях по-прежнему используют кинетические параметры «классического» уравнения Михаэлиса—Ментен — Vmax и Кт,каж.
Концентрация ионов в растворе — один из наиболее важных факторов, определяющих характер поступления ионов в корни. Растения, произрастающие при разной доступности иона в среде, адаптируются к условиям среды, изменяя не только морфологию растущих корней (см. рис. 6.4), но и качественные характеристики транспортных систем. Так, скорость поглощения К+ проростками ячменя, росшими при низкой концентрации К+ в среде (5 мкМ), возрастает почти в восемь раз, если в растворе, из которого шло поглощение в эксперименте, она составляла 100 мкМ (рис. 6.31).
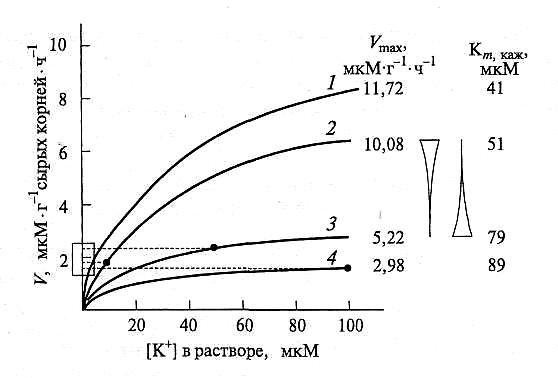
Рис. 6.31. Изменение скорости поступления ионов К+( F) в зависимости от их концентрации в растворе ([К+]) при кратковременной экспозиции 10-дневных проростков ячменя, предварительно выращенных в водной культуре с концентрацией К+, мкМ:
7-5; 2- 10; 3- 50; 4— 100.
Пунктирные линии от каждой кривой на ось ординат указывают на скорость поступления К в
условиях, когда его концентрация в экспериментальном растворе совпадала с концентрацией в
среде выращивания (по Siddiqi, Glass, 1983)
Проростки, росшие при более высокой концентрации, тоже реагировали на увеличение [К+] в «экспериментальном» растворе, но изменение скорости поступления было тем меньше, чем больше была концентрация иона в среде при выращивании. Оценка кинетических параметров свидетельствует, что системы транспорта К+ у корней проростков, росших при 5 мкМ, имеют наибольшую поглотительную способность (самая высокая Vmax) и наибольшее сродство к К+ (самая низкая Кт,каж) (рис. 6.31, величины Vmax и Кт даны справа от графиков).
У корней ячменя, росшего при более высоких концентрациях К+ в среде, Vmax снижается, а Кт,кaж возрастает. При этом скорости (V) поглощения К+ проростками разных групп при той концентрации, которая соответствуют условиям выращивания (рис. 6.31, экстраполяция на ось ординат), практически одинаковы и близки к величине 2 мкМ • г-1 • ч-1 (за исключением растений, росших при 5 мкМ). Следовательно, растения приспосабливаются поглощать «требуемое» количество К независимо от его концентрации в среде.
Таким образом, транспортные системы растения адаптируются к условиям произрастания, что обеспечивается синтезом и функционированием переносчиков и каналов с разными кинетическими характеристиками (сродством к иону и скоростями переноса). При поглощении ионов из почвенного раствора множество других факторов определяют зависимость «скорость поглощения — концентрация в среде»; такие, например, как перемешиваемость раствора, массовый ток, истощение в ризосфере, скорость диффузии ионов, процессы адсорбции на почвенных частицах и десорбции в раствор и т.д.