

Высоты поднятия воды (h) в капилляре в зависимости от его радиуса (г)
Объемный
поток воды для ксилемы вследствие этого
можно выразить как: Jw
= Lp∆P.
Согласно
закону Пуазейля, гидравлическая
проводимость (Lp)
цилиндрических
труб (такими в грубом приближении можно
считать элементы ксилемы) равна
![]() (ламинарное движение), где r
—
внутренний
радиус
трубы; η
—
вязкость раствора. Соответственно
объемный поток воды в
(ламинарное движение), где r
—
внутренний
радиус
трубы; η
—
вязкость раствора. Соответственно
объемный поток воды в
ксилемном элементе будет равен Jw = - ∆Р. Знак «минус» в уравнении
потока Пуазейля означает, что положительный объемный поток (Jw > 0) направлен в сторону уменьшения гидростатического давления (∆P< 0). Градиент гидростатического давления в элементах ксилемы и КС создается в основном благодаря транспирации. При интенсивной транспирации в ксилеме некоторых растений может развиваться большое натяжение (отрицательное гидростатическое давление) вплоть до -10 МПа, которое поддерживается за счет поверхностных явлений на границе ксилемный сок — стенка. Когда транспирация прекращается или ослабевает, что происходит ночью или при высокой влажности воздуха, гидростатическое давление в ксилеме падает до значений, близких к 0, а в некоторых случаях может принимать положительные значения (т. е. выше атмосферного). Экспериментально показано, что изменения гидростатического давления в ксилеме (от положительного до отрицательного) на протяжении суток приводят к заметным колебаниям в диаметре дерева. При положительном давлении вода выделяется из растения через гидатоды листьев (видоизмененные устьица).
Приведенное выше уравнение Пуазейля показывает, что диаметр проводящего воду канала имеет огромное влияние на его гидравлическую проводимость, а следовательно, и на величину потока воды. У растений поперечные размеры проводящих элементов ксилемы сильно колеблются. Например, около кончика корня кукурузы диаметр сосудов протоксилемы равен 5 — 10 мкм. На расстоянии 25 мм от кончика средний диаметр сосудов ранней метаксилемы составляет 23 мкм, а на расстоянии больше 250 мм от кончика сосуды поздней метаксилемы достигают в диаметре 100 мкм. Вследствие такого различия в диаметре сосуды поздней метаксилемы обладают значительно более высокой гидравлической проводимостью, чем сосуды ранней метаксилемы. Расчеты по уравнению Пуазейля показывают, что проводимость одного зрелого сосуда поздней метаксилемы с диаметром 100 мкм эквивалентна 357 сосудам ранней метаксилемы с диаметром 23 мкм. Созревание поздней метаксилемы имеет большое значение для зрелой части основного корня, который собирает воду, поступающую из многочисленных боковых корней, и транспортирует ее в стебель.
5.8.4. Движение воды в корне
Движение воды в радиальном направлении из почвенного раствора в проводящие элементы ксилемы осуществляется как непосредственно через клетки, так и по апопласту тканей корня (рис. 5.8). Напомним, что под термином апопласт у растений понимают единую систему клеточных стенок и межклеточных полостей. Транспорт воды через клетки получил название движение
воды от клетки к клетке («cell to cell»). В этом случае, преодолевая последовательно расположенные в радиальном направлении ткани корня, вода при движении от клетки к клетке может пересекать мембраны или идти по непрерывному цитоплазматическому пути через соединяющие соседние клетки плазмодесмы. Единая система, объединяющая с помощью плазмодесм протопласты растительных клеток, называется симпластом. В соответствии с этим путь движения воды от клетки к клетке подразделяется на трансмембранный и симпластный (рис. 5.8). При трансмембранном движении вода в пределах одной клетки дважды пересекает плазмалемму и клеточную стенку, а если заходит в вакуоль, то дважды пересекает и тонопласт. В трансмембранном движении воды важную роль играют образованные аквапоринами водные каналы, которые характеризуются высокой водопроницаемостью (см. подразд. 5.6). Таким образом, движение воды от клеток эпидермиса к сосудам ксилемы может осуществляться по трем параллельным направлениям: 1) через клеточные мембраны (трансмембранный путь); 2) по цитоплазме и плазмодесмам (симпластный путь); 3) по клеточным стенкам и межклетникам (апоп-ластный путь). Эти три пути движения воды не автономны: вода может перемещаться по комбинированным траекториям, например пройти некоторый путь по апопласту, затем пересечь ПМ и далее передвигаться от клетки к клетке по симпласту.

Рис. 5.8. Направления водного тока в растительных тканях (по Е. Steudle and С. A. Peterson, 1998)
В настоящее время отсутствуют экспериментальные подходы, которые позволили бы количественно оценить трансмембранный и симпластный потоки воды в корне. Тем не менее имеющиеся данные уже сейчас позволяют предположить, что при движении воды от клетки к клетке трансмембранные потоки значительно превосходят симпластные. Из-за отсутствия возможности различать транспорт воды через мембраны и по симпласту, как правило, используют упрощенную транспортную модель, в которой рассматривают два параллельных пути — путь от клетки к клетке, включающий потоки трансмембранного и симпластного путей, и апопластный путь. Оба потока существуют в растении одновременно. Вклад того или иного пути в суммарный поток воды определяется многими факторами, такими, например, как вид растения, его
возраст, условия выращивания, действие неблагоприятных факторов среды и т.д. Однако главным фактором, определяющим относительный вклад клеточного и апопластного путей в общий поток воды в корне, является интенсивность транспирации.
При апопластном транспорте от поверхности клеток корня до просвета в элементах ксилемы и далее в восходящем направлении в побеги вода ни разу не пересекает мембраны, а сам апопласт не обладает свойством полупроницаемости. Все это означает, что осмотические явления не вовлечены в формирование движущих сил апопластного транспорта воды. В этой связи ток воды через апопласт имеет гидравлическое происхождение, так как движущей силой для него служит преимущественно градиент гидростатического давления, и ток воды по апопластному пути также называют гидравлическим. Основным процессом, приводящим к формированию ∆Р, при апопластном (гидравлическом) транспорте является транспирация, что и определяет ее влияние на соотношение потоков воды через клетки и апопласт. При отсутствии водного дефицита в дневное время, когда устьица открыты и интенсивность транспирации высока, поток воды через апопласт в значительной степени превышает поток через клетки. При низкой интенсивности транспирации преобладающим становится движение воды от клетки к клетке.
При движении воды от клетки к клетке, когда вода на своем пути пересекает мембраны, явления осмоса играют определяющую роль. Ток воды в этом случае называют осмотическим, хотя градиент водного потенциала для него включает не только осмотическую, но и гидравлическую составляющую. Градиент водного потенциала между двумя клетками описывается уравнением
∆ψ = ∆Р - σ∆π, (5.16)
где ∆Р, ∆π — разности тургорного и осмотического давления между двумя клетками соответственно; σ — коэффициент отражения мембран. С учетом того, что σ для мембран стремится к единице, выражение для ∆ψ принимает вид
∆ψ = ∆Р - ∆π.
Следует обратить внимание на то, что при апопластном транспорте воды и транспорте воды через клетки генерация ∆Р обусловлена в целом разными процессами. Как обсуждалось выше, апопластный транспорт связан в основном с градиентом гидростатического давления, создаваемым натяжением жидкости в проводящих элементах ксилемы. Основной вклад в ∆Р при транспорте воды через клетки вносит градиент тургорного давления, развивающегося в клетках корня.
В условиях пониженной транспирации поток воды через клетки преобладает над ее потоком через апопласт. Поскольку клеточные мембраны высокопроницаемы для воды, градиент водного потенциала между клетками и апопластом быстро уравновешивается, создавая локальные равновесия между двумя указанными потоками.
Так как для апопласта коэффициент отражения стремится к нулю, а для мембран — близок к единице, коэффициент отражения целого корня составляет некоторую промежуточную величину. Если вышеприведенное уравнение (5.16) использовать для целого корня, в котором вода движется как по апопласту, так и через клетки, то оно примет вид
∆ψ
=
∆Р
- σ![]() ∆π,
(5.17)
∆π,
(5.17)
где ∆ψ, ∆Р, ∆π — градиенты соответственно водного потенциала, гидростатического давления и осмотического давления между почвой и ксилемным соком в корне; σ — коэффициент отражения корня. Величина σ ∆π в приведенном уравнении выражает эффективную составляющую осмотического градиента водного потенциала при движении воды через целый корень. Экспериментальное определение коэффициента отражения показывает, что для целого корня эта величина изменяется в пределах 0,2 — 0,8.
Поддержание необходимых концентраций осмотически активных частиц в клетках и формирование ∆Р и ∆π в радиальном направлении корня, вызывающее движение воды от клетки к клетке и далее к проводящим элементам ксилемы, осуществляется за счет (7) поглощения ионов клетками корней из почвы и (2) биосинтеза в цитоплазме клеток низкомолекулярных органических соединений, которые получили название осмолитов (см. подразд. 8.2.2). К основным функциям осмолитов относятся поддержание осмотического равновесия между внутриклеточными компартментами и осморегуляция клеток при водном дефиците (см. подразд. 8.2.2).
Поглощение ионов из почвы и биосинтез осмолитов в клетках корня могут обеспечить движение воды от клетки к клетке в радиальном направлении в отсутствие транспирации или при ее очень низкой интенсивности, т. е. когда гидравлический поток по апопласту мал. В этом случае движение воды к элементам ксилемы обусловлено процессами, протекающими преимущественно в клетках корня. Силы, под действием которых вода перемещается из почвенного раствора в сосуды ксилемы корня в отсутствие транспирации, называют корневым давлением, а механизм, который его осуществляет, — нижним концевым двигателем транспорта воды.
Корневое давление можно наблюдать при удалении надземной части растений. Истечение из перерезанных сосудов ксилемы сока, получившего название пасоки, является результатом транспорта воды из почвы в ксилему, который осуществляется без участия надземных органов и, следовательно, транспирации. Корневое давление проявляется в весеннем движении сока у березы и клена, когда листья на растениях еще не сформировались, т. е. транспирация практически отсутствует. Упоминавшийся выше процесс гуттации, наблюдаемый при закрытых устьицах, также возникает вследствие корневого давления. Считается, что корневое давление создается за счет транспорта воды по градиенту водного потенциала. При этом предполагается наличие такого процесса, как загрузка ионами или другими веществами ксилемы корня, — процесса приводящего к повышению осмотического давления ксилемного сока (понижению его водного потенциала) и формированию AT, направленного из почвы в ксилему.
Вода, двигаясь в радиальном направлении корня к проводящим элементам ксилемы, проходит через разные последовательно расположенные ткани: эпидермис, кору, эндодерму, перицикл, стелярную паренхиму и стенки сосудов (рис. 5.9). Поэтому ток воды в радиальном направлении корня можно рассматривать как движение воды в гидравлической цепи через разные по величине и последовательно расположенные гидравлические сопротивления. Рассмотрим путь воды в радиальном направлении по апопласту корня с учетом гидравлической проводимости разных типов тканей и начнем со стенок сосудов.
Вода, чтобы попасть в просвет сосудов, должна пройти через их стенки. Полагают, что основной ток воды идет через окаймленные поры, а не через утолщенные и лигнифицированные части стенок. В корнях кукурузы такие поры имеют диаметр 2 мкм и занимают до 14 % от общей площади поверхности сосудов ранней метаксилемы. Вклад стенок сосудов в общее сопротивление транспорту воды в корне кукурузы оценен экспериментально. Показано, что стенки сосудов отвечают не более чем за 10 —30 % общего сопротивления движению воды в радиальном направлении. У других видов растений эта величина может быть еще меньше.
Апопласт эндодермы корня включает пояски Каспари (рис. 5.9).
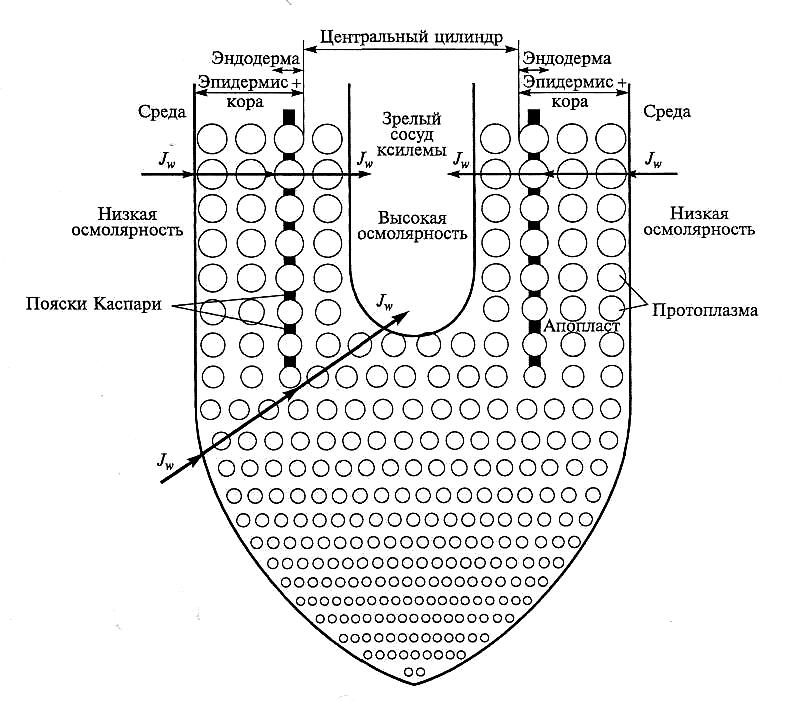
Рис. 5.9. Транспортная модель корня (пояснение в тексте) (по E.Steudle and C.A.Peterson, 1998)
Последние представляют собой отложения лигнина, а в некоторых случаях и суберина. До недавнего времени полагали, что движение воды по апопласту в радиальном направлении корня прерывается на уровне эндодермы, так как пояски Каспари представляют для нее непреодолимую преграду. В соответствии с этой точкой зрения вода и растворенные в ней вещества, достигнув по апопласту эндодермы, должны перейти в симпласт, чтобы продолжить свое движение в радиальном направлении корня. Если основной ток воды осуществляется по апопласту, а пояски Каспари — главный барьер на пути ее движения, то повреждение эндодермы должно значительно увеличить гидравлическую проводимость корня. Было, однако, показано, что в молодых зонах корня кукурузы гидравлическая проводимость заметно не увеличивается при повреждении эндодермы. Результаты этих опытов привели к заключению, что эндодерма с ее поясками Каспари не может быть главным препятствием на пути движения воды к сосудам ксилемы.
Сделанный вывод был подтвержден биохимическими исследованиями, которые показали, что основным химическим компонентом поясков Каспари является не суберин, как полагали ранее, а лигнин. Поскольку в отличие от суберина лигнин — гидрофильное соединение, содержащие его структуры не могут быть препятствием на пути движения воды. Если пояски Каспари содержат не только лигнин, но и суберин, который встречается в КС корня у многих видов растений, то гидравлическая проводимость может заметно снизиться. Таким образом, по крайней мере, у молодых корней стенки проводящих элементов ксилемы и эндодерма без субериновых отложений не создают существенного препятствия на пути водного тока. Следует подчеркнуть, что при относительно низком сопротивлении апопласта эндодермы водному току апопласт является препятствием для движения ионов. При всех условиях транспорт ионов по апопласту корня прерывается на уровне эндодермы, и они переходят в симпласт, чтобы продолжить свое движение в радиальном направлении к сосудам ксилемы.
В экспериментах на молодых корнях кукурузы было показано, что повреждение эндодермы приводит к резкому падению корневого давления, которое свидетельствует о выходе ксилемного раствора через повреждение из компартмента с высоким осмотическим давлением (центральный цилиндр) в компартмент с низким давлением (внешняя среда). Эти результаты подтвердили давно предполагавшуюся роль эндодермы как барьера для обратной диффузии ионов из центрального цилиндра в апопласт тканей коры. Эндодерма удерживает ионы в центральном цилиндре, обеспечивая, с одной стороны, высокую эффективность снабжения надземных органов элементами минерального питания, а с другой — формирование корневого давления. Таким образом, транспортные свойства корня оптимизированы как для воды, так и для ионов. Поглощенные корнем ионы удерживаются в центральном цилиндре, в то время как низкое гидравлическое сопротивление эндодермы позволяет в достаточной мере обеспечивать водой надземные органы.
Движение воды от поверхности корня к эндодерме лежит через клетки эпидермиса и коры. Исследование гидравлической проводимости этих тканей показало, что в корне главное сопротивление водному току оказывают ткани коры. Удаление некоторой части коры уменьшает его приблизительно пропорционально числу удаленных слоев этой ткани.
Выше, при рассмотрении тканей, через которые вода движется от периферии корня к центру, основное внимание было сфокусировано на апопластном пути. Однако путь от клетки к клетке играет не менее важную роль в транспорте воды к проводящим элементам ксилемы. У всех растений такой путь воды от клетки к клетке доминирует в отсутствие градиента гидростатического давления, создаваемого транспирацией. У некоторых видов, например у гороха (Pisum sativum) и ячменя (Hordeum vulgare), путь от клетки к клетке преобладает при всех условиях.