

5.8. Движение воды в целом растении
Вода образует непрерывную фазу в системе почва—корень—стебель—лист— атмосфера. Поглощенная корнями из почвы вода транспортируется по стеблю в листья. Лишь незначительная доля всей поглощенной воды потребляется клетками растения. Подавляющая часть воды, оказавшейся в растении, достигнув листьев, испаряется. В глобальном масштабе растения возвращают обратно в атмосферу до половины того количества воды, которое ежегодно выпадает на Землю в виде осадков.
5.8.1. Теория когезии и натяжения
Более 100 лет назад Дж. Бём разработал теорию когезии и натяжения, которая легла в основу современной модели транспорта воды в целом растении. В середине прошлого столетия Т.Х.Ван ден Хонерт, основываясь на физико-химических свойствах воды, в рамках этой теории разработал термодинамические принципы дальнего транспорта воды (транспорт между органами растений и в целом растении). Основные положения теории когезии и натяжения формулируются следующим образом.
1. За счет водородных связей между молекулами воды возникают большие силы сцепления (когезии). В связи с этим вода может подвергаться натяжению величиной от десятков до нескольких сот МПа, прежде чем столб воды, находящейся в капилляре, в частности в сосуде ксилемы, разрушится.
2. Вода в растениях формирует непрерывную водную систему, идущую от почвы к поглощающей поверхности корня и далее через корень и стебель к испаряющей поверхности листьев. За некоторым исключением, ток воды в этой непрерывной системе является гидравлическим по происхождению, и система может быть описана как цепь сопротивлений, размещенных последовательно и параллельно.
3. Испарение воды листьями приводит к снижению их водного потенциала и тем самым заставляет воду двигаться из ксилемы к испаряющим клеткам листьев. Это в свою очередь снижает давление в ксилеме до значений, меньших атмосферного давления (Р < 0).
4. Вдоль транспирирующего растения устанавливается градиент давления. Это является причиной тока воды из почвы в корни и далее через стебли к испаряющим воду листьям. Разность в водном потенциале между атмосферой и почвой достаточна для подъема воды даже у самых высоких деревьев.
5. Когда натяжение в сосудах ксилемы достигает некоторой критической величины, через поры в стенках сосудов проникают пузырьки воздуха. Объединяясь, они могут сформировать пузырек достаточной величины, чтобы разорвать столб ксилемного сока в сосуде, т. е. привести к эмболии.
Рассмотрим в общих чертах ток воды в растении, приняв за основу вышеприведенные положения.
По аналогии с законом Ома поток воды (Jv) в гидравлической цепи представляют как произведение гидравлической проводимости (Lp — величина, обратная гидравлическому сопротивлению R) на градиент водного потенциала (Jv = Lp∆ψ). В соответствии с этой аналогией, если цепь состоит из нескольких сопротивлений, расположенных последовательно, в цепи течет гидравлический ток, величина которого может быть определена уравнением
Jv= ,
(5.14.)
,
(5.14.)
где Jv — поток воды через гидравлическую цепь; ∆ψi — разность водных потенциалов между концами i-го элемента цепи; Ri — сопротивление элемента i; R — сопротивление всей цепи. Если элементы соединены параллельно, суммарный поток воды может быть описан следующим образом: Ji = ∑Li ∆ψi = Lp∆ψ, где Li — гидравлическая проводимость i-го элемента цепи; Lp — гидравличеcкая проводимость всей цепи; ∆ψ — разность водных потенциалов между концами цепи. В гидравлической цепи целого растения выделяют три основных последовательно соединенных сопротивления: (R1) сопротивление на участке от поверхности клеток эпидермиса до сосудов ксилемы; (R2) сопротивление, которое оказывают элементы ксилемы; (R3) сопротивление при испарении воды в атмосферу из листьев (рис. 5.6). В цепи целого растения ксилема характеризуется наименьшим сопротивлением, тогда как самое большое сопротивление возникает при диффузии воды из листьев в атмосферу. В каждом из трех участков цепи целого растения можно выделить более дробные элементы, соединенные между собой последовательно или параллельно. Последовательно соединены ткани, которые вода пересекает, передвигаясь от периферии корня к центру; параллельно соединенные элементы — апопластный и симпластный пути движения воды.
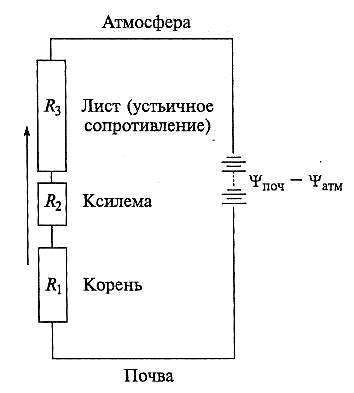
Рис. 5.6. Упрощенная модель установившегося тока воды по растению. Стрелка указывает направление движения воды в системе почва— растение — атмосфера
Испарение воды с поверхности листьев в атмосферу определяет направление движения воды во всей системе почва—корень—стебель—лист—атмосфера и генерирует движущие силы для поглощения воды корнями. Снижение водного потенциала листьев при транспирации увеличивает градиент водного потенциала на участке ксилема — листовая пластинка, что приводит к усилению поглощения воды клетками листа из ксилемы и соответственно к стимуляции поглощения воды корнями из почвы. Падение водного потенциала на каждом участке системы происходит в соответствии с величиной его сопротивления. Рассмотрим весь путь движения воды по отдельным элементам гидравлической цепи. Ради удобства изложения этапы водного пути будем рассматривать в обратном порядке, т. е. от атмосферы к почве.