

Триптофановий оперон – приклад репресибельних оперонів
У триптофановом опероні, як і в лактозном, теж є подвійний механізм регуляції. По-перше, як завжди, регулюється переміщення РНК-полімерази по оператору. Другим же (і чутливішим) об'єктом регуляції є не зв’язування РНК полимерази з промотором, а закінчення транскрипції на атенюаторі.
Нагадаємо: атенюатор - це присутня в деяких оперонах ділянка ДНК між оператором і генами, на якому за певних умов припиняється транскрипція оперону.
Як правило, оперони, що мають атенюатор, є репресибельними і контролюють синтез (анаболізм) того або іншого необхідного компонента - наприклад, аминокислоти: триптофану, гістидину, фенілаланина.
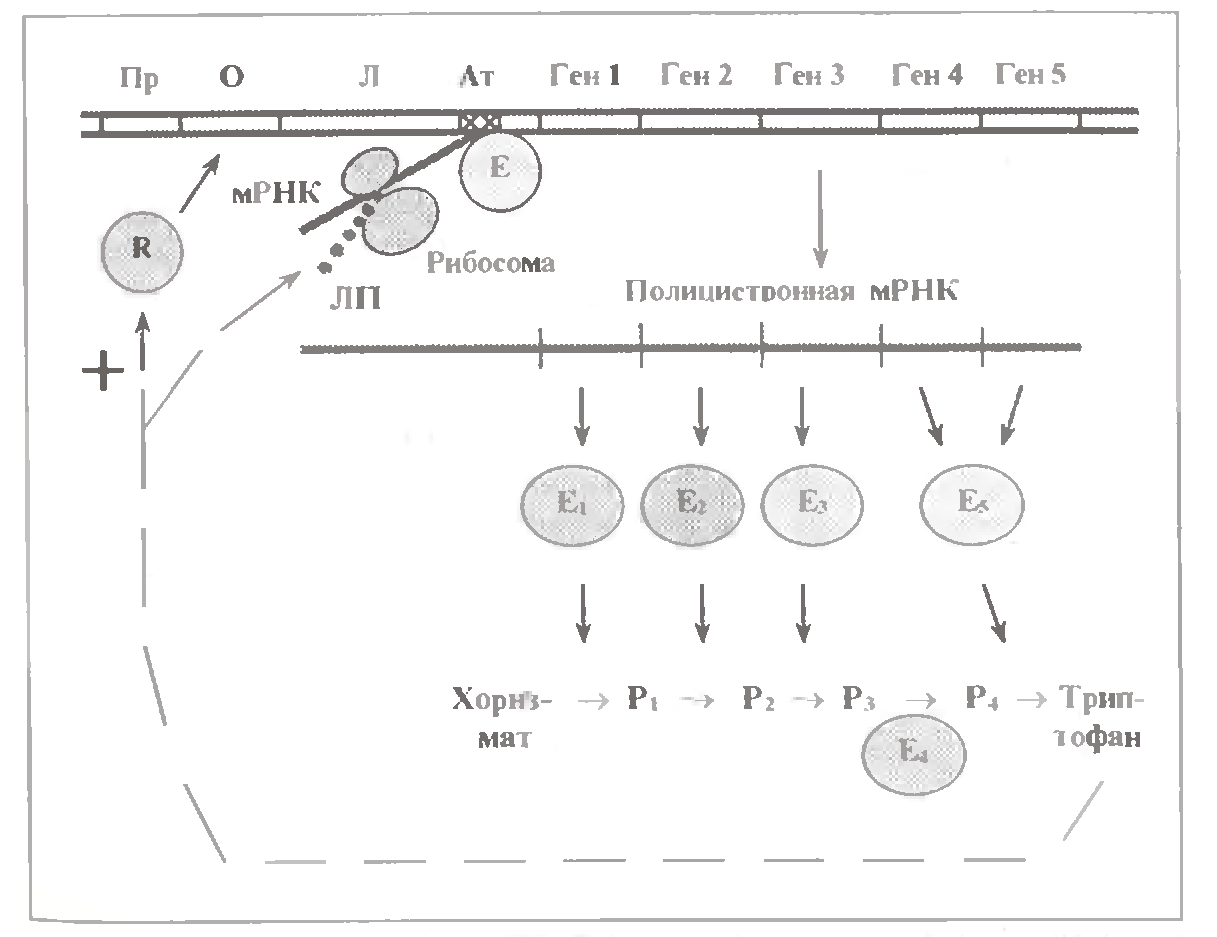
Схема триптофанового оперону: Пр – промотор, О - оперон, Л – лидерний відділ оперона, Ат – атенюатор, R – білок репресор, Е- РНК-полімераза, ЛП – лідерний пептид, Е1…Е5 – ферменти синтезу триптофана, Р1…Р4 – проміжні метаболіти синтезу триптофана.
Триптофановый оперон включає 5 цистронів які кодують 4 ферменти заключного етапу утворення триптофану. При цьому останній фермент містить субодиниці 2 видів, тому кодується двома цистронами. Разом з тим, ген останнього ферменту ланцюга знаходиться поза даним опероном.
За промотором і оператором в оперонах цього типу знаходиться т. з. лідерний відділ; саме він закінчується атенюатором.
In процесі транскрипції цього відділу утворюється лідерна ділянка мРНК. Останній тут зв'язує рибосому і починаэ трансляцію з утворенням лідерного пептиду (ЛП). Ключова особливість останнього - серед його 14 амінокислотних залишків містяться 2 залишки триптофану, тобто тої самої амінокислоти, синтез якої контролюється опероном.
Аналогічно, в ЛП фенілаланінового оперону серед 15 залишків - 7 залишків фенілаланіна, а в ЛП гістидинового оперона - 7 підряд залишків гістидину. Тому механізм атенюаторної регуляції в усіх цих випадках однаковий.
Повернемося до триптофановому оперону. Коли in клітині достатня кількість триптофану, то синтез лідерного пептиду йде без затримки: рибосома, що його утворює, відстає від РНК-полімерази. В цих умовах при досягненні РНК-полімеразою атенюатора з високою часткою ймовірності спрацьовує сигнал про закінчення транскрипції: РНК-полімераза диссоціює на ДНК і гени не зчитуються.
Таким чином, триптофан, швидко включаючись в лідерний пептид, блокує через атенюаторний механізм синтез ферментів, необхідних для його утворення. Правда, блокування це - не повне, оскільки зберігається невелика вірогідність того, що РНК-полімераза все ж здолає атенюаторну ділянку.
Якщо, навпаки, концентрація триптофану в клітині низька, то рибосома затримується з синтезом лідерного пептиду і відстає від РНК-полімерази. Це так змінює конфігурацію ДНК або лідерного відділу мРНК, що сигнал про закінчення трансляції на атенюаторі не спрацьовує. Кожна молекула РНК-полімерази проходить цю "небезпечну" ділянку і транскрибує гени. Тобто активно синтезуються ферменти, необхідні для поповнення запасу триптофану в клітині.
Відмітимо: зміст триптофану в самих цих ферментах дуже невелике. Тому його дефіцит в клітині не дуже сильно позначається на швидкості трансляції структурних цистронів мРНК (на відміну від синтезу лідерного пептиду).
Отже, через атенюаторный механізм триптофан може пригнічувати активність "свого" оперону, але, як було сказано, не до кінця.
Глибше пригнічення активності оперону відбувається при дуже високій концентрації триптофану. Тоді реалізується другий (і загальний) спосіб регуляції. Триптофан зв’язуються із специфічним білком-репресором і підвищує його спорідненість до оператора триптофанового оперону. Це повністю блокує цей оперон.