

Генетичний код
Говорячи про код, досі ми мали на увазі змістовний ланцюг ДНК. Але така ж, з урахуванням заміни тиміну (Т) на урацил (У), послідовність нуклеотидів в про-мРНК (чи, з деякими обмовками, мРНК).
Триплети мРНК, відповідні триплетам ДНК, називаются кодонами. Дійсно, саме вони безпосередньо визначають порядок включення амінокислот в пептидний ланцюг, що синтезується на рибосомі.
З тієї ж причини в таблиці генетичного коду (розшифрованого в ході модельних експериментів) завжди вказують не триплети змістовного ланцюга ДНК, а кодони мРНК.
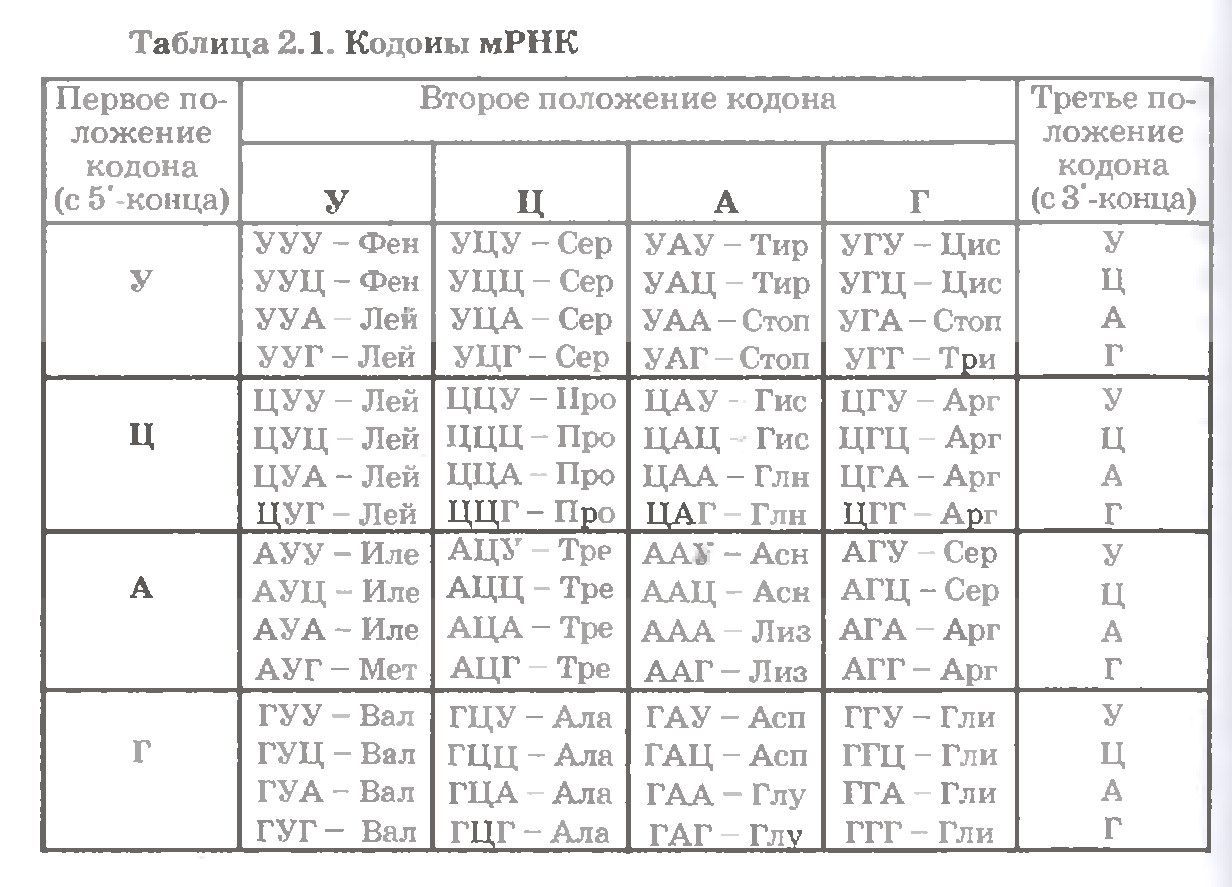
З цієї таблиці видно, що код, дійсно, є виродженим: для усіх амінокислот, окрім метіоніну і триптофана, є по 2 кодони і більше. У тому числі по 6 кодонів для трьох амінокислот - аргініну, лейцину і серину.
Крім того, можна зробити ще два заключення.
а) В більшості випадків кодони однієї амінокислоти розрізняються лише останнім (третім) нуклеотидом.
Відмінності ж по другому або першому нуклеотиду спостерігаються лише тоді, коли амінокислоті відповідає більше 4 кодонів, тобто коли варіації по третьому нуклеотиду вичерпані.
б) у схожих по будові амінокислот кодони також схожі між собою: співпадають після двох нуклеотидів або (рідше) по одному, але центральному, нуклеотиду.
Наприклад, кодони гомологів - Асп і Глу - співпадають по перших двом нуклеотидам; кодони Глу і його амида - Глн - по двох останнім; кодони 5 амінокислот з неполярним радикалом (Фен, Лей, Ілей, Мет, Вал) - по центральному нуклеотиду. І так далі.
Скорочені позначення амінокислот : Ала - аланін, Арг - аргінін, Асн - аспарагін, Асп аспарагінова кислота, Вал - валін, Гіс - гістидин, Глі - гліцин, Глн - глутамін, Глу - глутаміновая кислота, Ілей - ізолейцин, Лей - лейцин, Ліз – лізин, Мет - метіонін. Про - пролін, Сер – серин, Тир - тирозин, Тре – треонін, Три – триптофан, Фен – фенілаланін, Цис -цистеїн.
Стоп -"безглуздий" кодон, що перериває трансляцію мРНК.
Можливо, це пояснюється тим, що спочатку (на ранніх стадіях еволюції) були кодони лише для невеликої групи перших амінокислот. Потім розширення великої кількості амінокислот шляхом їх модифікації супроводжувалося модифікацією і початкових кодонів, тобто заміною одного, рідше двох, нуклеотидів. Але інші два нуклеотиди (чи один центральний) залишалися колишніми.
Познайомившись з функціональними елементами ДНК і способом запису генетичної інформації, продовжимо розгляд цих елементів.
Найцікавіше питання - як вони взаємодіють один з одним, тобто які складніші об'єднання утворюють і як останні функціонують за тих або інших умов.
По суті справи, питання про організацію генетичного матеріалу найтіснішим чином пов'язаний з ключовим питанням про регуляції активних генів.
Найкраще ці аспекти вивчені у бактерій.
Потім перейдемо до генетичного матеріалу еукариот.
Оперонная організація генетичного матеріалу у бактерій Регуляторні і конститутивні гени Загальна схема оперону
У бактерій гени ферментів, що каталізують ряд послідовних реакцій, нерідко об'єднуються в одну структурно-функціональну одиницю - оперон.
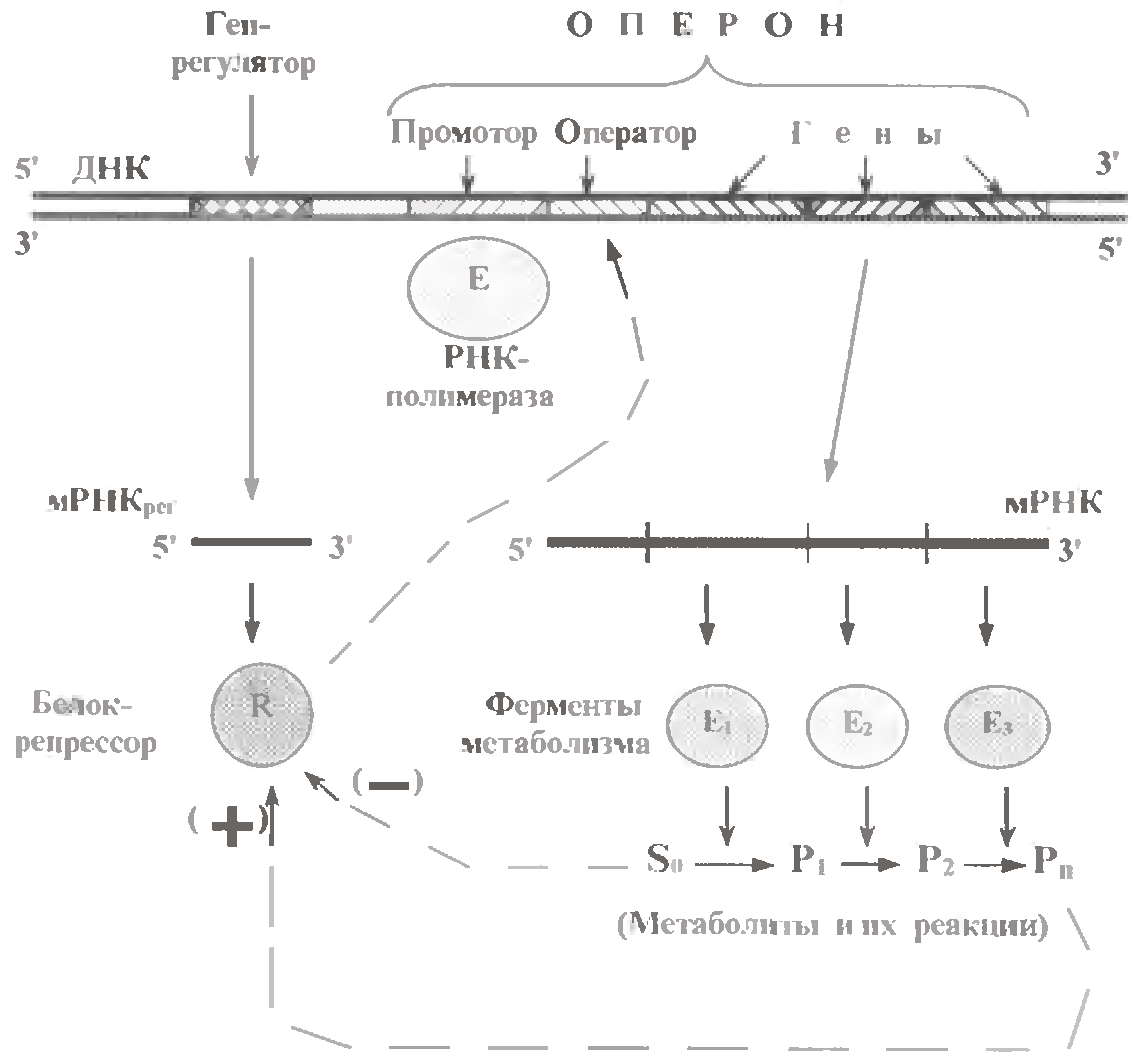
Окрім вказаних генів, в оперон входять промотор (місце зв'язування РНК-полімерази) і оператор (місце зв’язування білка-репресора).
Сам же білок-репресор кодується спеціальним геном-регулятором, який до складу оперону зазвичай не входить. Для різних оперонів існують різні білки-репресори і, відповідно, різні гени-регулятори. Хоча не виключено, що якісь білки-репресори контролюють активність відразу декількох оперонів.
Принцип регуляції активності оперону полягає в тому, що на спорідненість білка-репресора до оператора можуть впливати метаболіти того ланцюга реакцій, ферменти якої кодуються данним опероном.
У зв'язку з цим розрізняють оперони двох типів.
а) Індуцибельні оперони:
регулятором є початковий субстрат (S0) ланцюга контролюємих реакцій;
у відсутність цього субстрату білок-репресор має високу спорідненість до оператора, тому РНК-полімераза не може транскрибувати гени оперону (оперон "Виключений");
при накопиченні метаболита S0 в клітині, деяка кількість його зв'язується з білком-репресором, знижуючи спорідненість останнього до оператора; оперон "включається" - і синтезуються ферменти, що забезпечують перетворення речовини S0.
Як видно, регуляція оперону речовиною S0 це приклад прямого позитивного зв'язку : початковий субстрат стимулює кінець кінцем реакції свого метаболізму.
б) Репрессибельні оперони:
регулятором служить кінцевий продукт (Рn) ланцюга контрольованих реакцій;
у відсутність цього продукту білок-репресор має низьку спорідненість до оператора, тому РНК-полімераза транскрибує гени оперону - оперон "включений", і синтезуються ферменти, сприяючи утворенню речовини Рn;
при накопиченні ж цієї речовини деяка його кількість зв'язується з білком-репресором і підвищує спорідненість останнього до оператора - оперон "вимикається", синтез відповідних ферментів і утворення метаболіту Рn припиняється.
Тут, на відміну від попереднього випадку, ми бачимо регуляцію за типом негативного зворотного зв'язку: метаболіт гальмує у результаті реакції, що ведуть до його утворення.
Такі загальні схеми функціонування індуцибельних і репресибельних оперонів. Але організація конкретних оперонів нерідко включає ті або інші додаткові деталі які іноді істотно ускладнюють картину. Два таких конкретних оперона ми розглянемо трохи нижче.