Руйнування мРнк еукаріот із 3'-кінця Роль пів(а) -фрагмента
У еукаріот тривалість існування мРНК в клетині (точніше, час напівжиття) істотно більше від 10 хвилин (для короткоживучих мРНК) до 2 діб. При цьому короткоживучими зазвичай бувають мРНК регуляторних білків - для того, щоб при зміні якихось параметрів клітина могла швидко відреагувати зміною синтезу цих білків.
Друга особливість обумовлена тим, що мРНК еукаріот - моноцистронні. Тому ефекту положення, подібного вищеописаному, тут бути не може, навіть якщо б мРНК зруйнувалась з 5'- кінця.
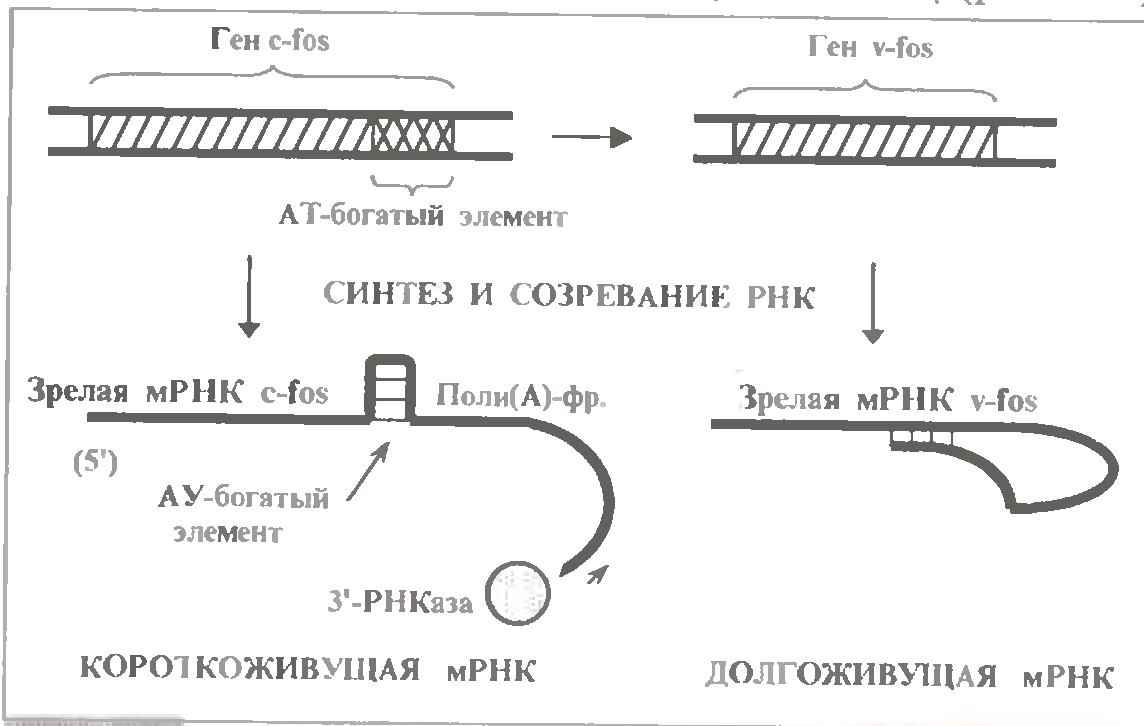
Але є і третя особливість: розпад мРНК у еукаріот здійснюється не 5´-РНКазами, а 3'-РНКазами (і, можливо, ендонуклеазами). Тобто зазвичай він починається з того кінця, де практично усі мРНК (окрім гістонових) містять пів(А) -фрагмент з приблизно 200 нуклеотидів.
Отже, подібно до теломер ДНК, цей фрагмент служить свого роду буферною зоною, поступове укорочення якого захищає до певного часу від такої ж долі кодучу частину нуклеїнової кислоти.
Причому, згідно двічі згадуваній гіпотезі "білетиків", пів(А) -фрагмент руйнується 3'-РНКазой не постійно, а періодично: в тісному зв'язку з функціонуванням мРНК. Потім завершения трансляції мРНК черговою рибосомою від пів(А) фрагмента відщепляється 10-15 нуклеотидів. Коли ж в цьому фрагменті залишається всього біля 50 нуклеотидів, мРНК стає доступною для РНКаз і швидко руйнується.
Звідси витікає, що звичайна кратність трансляції мРНК повинна складати 10-15 разів. Наприклад, втративши 15 разів по 10 нуклеотидів, пів(А) фрагмент коротшатиме з 200 нуклеотидів до критичної довжини в 50 нуклеотидів.
Як пояснити такий зв'язок розпаду пів(А) -фрагмента з трансляцією? Можливий механізм показаний на мал.
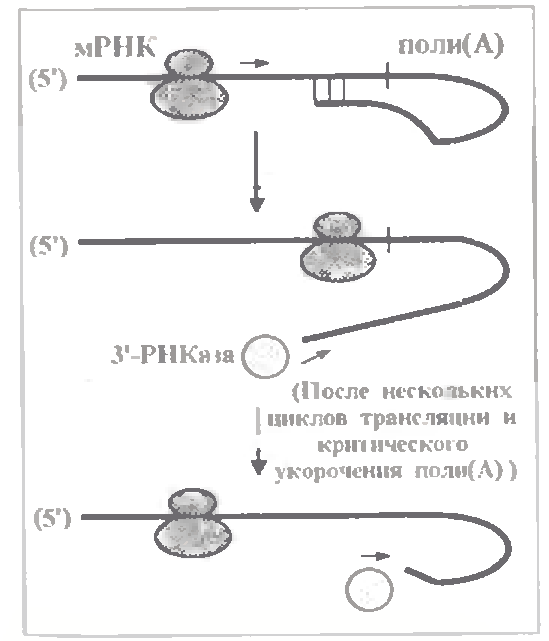
Згідно з ним, пів(А) -фрагмент утворює петлю за рахунок взаємодії своїм 3´-кінцем з певною ділянкою трансльованої частини мРНК. І в такій дволанцюговій структурі 3' -кінець недоступний для 3'-РНКази.
Коли ж по вказаній ділянці проходить рибосома, петля на якийсь час розривається і 3'-кінець стає доступним для РНКази, яка устигає по черзі відщепнути 10-15 нуклеотидів. Потім (при звільненні ділянки від рибосоми) петля знову відновлюється, але вже укорочена.
Нарешті, коли від пів(А)~фрагмента залишається всього 50 нуклеотидів, петля утворитися не може і РНКаза без перешкод руйнує увесь ланцюг мРНК.
Роль ау-элементів
А з чим пов'язані значні відмінності в тривалості життя різних мРНК?
Виявляється, у короткоживучих мРНК в 3'- нетрансльованій області (розташованої між кодуючою частиною і пів(А) -фрагментом) знаходяться спеціальні АУ-збагачені елементи (AYRE - AU - rich elements). Їх присутність різка пришвидшує деградацію пів(А) -фрагмента.
Можливо, такі елементи перешкоджають утворенню пів(А) -фрагментом петлі, що захищає 3´-кінец (мал.).
Цікавим прикладом, що ілюструє роль АУ-елементів, є мРНК гена c-fos. Останній - один з протоонкогенів, дуже схожий з вірусним онкоген ϑ- fos. (Приставка с- означає клітинний (cellular) ген, а приставка ϑ-вірусний ген). Обидва гени кодують один з чинників транскрипцій.
Так от, в мРНК гена з c-fos є АУ-елемент, і ця мРНК є короткоживучою. У складі ж мРНК онкогена ϑ- fos такого елементу немає; відповідно, час її напівжиття істотно більше, ніж, очевидно, і обумовлено виникнення пухлини. А для перетворення протоонкогена на онкоген вимагає лише делеція невеликої області (відповідною АУ-елементові мРНК).