Гормональная регуляция уровня глюкозы в крови. Гипергликемические гормоны: адреналин, глюкагон, глюкокортикоиды. Гипогликемические: инсулин.
Глюкагон. Образуется в альфа клетках островков Лангерганса поджелудочной железы. Полипептид (29 аминокислотных остатков). Образуется из проглюкагона путём ограниченного протеолиза.
Клетки мишени – печень, жировая ткань, мышцы (чутка).
Действие на углеводный обмен противоположно инсулину (просто поменяйте местами слова стимулирует и подавляет).
Глюкокортикоиды – гормоны, которые выделяются пучковой зоной коры надпочечников, из которых наиболее известен кортизол. О нём и поговорим.
Стероидной природы (холестерол – прегненолон – прогестерон – кортизол).
Ткани мишени – почти все ткани, в основном – мышцы, жировая ткань, печень, почки, лимфатическая ткань. Выделяется в ответ на андренокортикотропный гормон, который выделяется в ответ на кортиколиберин, который выделяется в ответ на стресс.
Транспортная форма – транскортин
Стимулирует глюконеогенез (фосфоенолпируваткарбоксиназы)
Стимулирует распад гликогена (гликогенфосфорилазу)
Адреналин: Стимулирует распад гликогена (гликогенфосфорилазу)
Концентрация глюкозы как интегральный показатель углеводного обмена в организме.
Все углеводы, которые поступают к нам с пищей распадаются либо до глюкозы, либо до других моносахаридов, которые в печени в итоге переходят в глюкозу. Все пути метаболизма глюкозы содержат глюкозу как исходный или как конечный субстрат. Это всё позволяет нам определить глюкозу как главный показатель обмена углеводов.
Глюкозо-6-фосфат – центральный метаболит внутриклеточного обмена глюкозы: как только глюкоза проникает в ткани она сразу же фосфорилируется в глюкозо-6-фосфат под воздействием глюкокиназы (в печени), гексокиназы (во всех остальных клетках). Это обусловлено несколькими причинами: 1. Фосфорилированная глюкоза уже не может выйти из клетки. 2. Поскольку глюкоза переходит в глюкозо-6-фосфат, то количество глюкозы в клетке падает, что создаёт хороший градиент для диффузии глюкозы в клетку. 3. Реакция превращения глюкозы в глюкозо-6-фосфат ингибируется АТФ. Значит, если у нас много АТФ, то энергия как бы и не нужна, тогда концентрация глюкозы в клетке и в крови выравнивается и глюкоза перестаёт поступать в клетку.
Возможные причины гипергликемии: переел глюкозы, диабет обоих типов, избыток глюкагона или глюкокортикоидов.
Гипогликемии: голодание, недостаток глюкагона, избыток инсулина, физические нагрузки интенсивные.
Нарушения углеводного обмена при сахарном диабете: поскольку углеводы не депонируются в ткани, то нарушается синтез гликогена в печени и мышцах, а также замедляется их переход в жиры. На самом деле, поскольку глюкоза не попадает в ткани, страдают любые пути ката- или анаболизма углеводов.
Механизм развития нарушений: есть два типа диабета – инсулинзависимый – когда нарушена выработка инсулина бета клетками островков Лангерганса поджелудочной железы (воспаления, аутоимунные реакции), и инулиннезависимый – когда инсулин нормально вырабатывается, но его воздействия на клетку нарушено.(дефекты рецепторов)
Лабораторные показатели: Тест толерантности к глюкозе, определение гликозилированного гемоглобина, определение количества инсулина и С-пептида в моче, по количеству альбумина в моче (альбуминурии), определение количества кетоновых тел.
Неэнзиматическая гликация – неферментное, ковалентное встраивание глюкозы или фруктозы в структуру мембраны белков при гипергликемии. В норме происходит в ничтожно малых количествах, при гипергликемии гликации подвергается всё и вся.
Роль в механизме развития осложнений гипергликемии: образуются гликозилированный гемоглобин (в принципе это не особо беда, но гемоглобин уже свою функцию не выполняет), гликозилированные кристаллины (белки хрусталика, из-за этого возникает катаракта), гликозилирование белков мембран сосудов, из-за чего развиваются ангиопатии, нефропатии и ретинопатии.
Клинико-диагностическое значение исследования фруктозамина и гликозилированного гемоглобина: ну, раз гликозилирование белков наблюдается при гипергликемии, значит мы может диагностировать гипергликемию определяя фруктозами (гликозилированный альбумин) и гликозилированный гемоглобин в крови. Причём гемоглобин живёт от 90 до 120 дней. Значит у нас уже 3 месяца как может и не быть гипергликемии, а гемоглобин останется. Про это любят задавать вопрос, так что будьте во всеоружии.
Чё это? Становление процессов переваривания в онтогенезе… Врождённая недостаточность…. Обмен глюкозамингликанов… А-а-а, для студентов педиатрического факультета! Ну что ж чуваки, облом, не судьба, не повезло, не фортануло.
Классификация, строение и биологическая роль липидов.
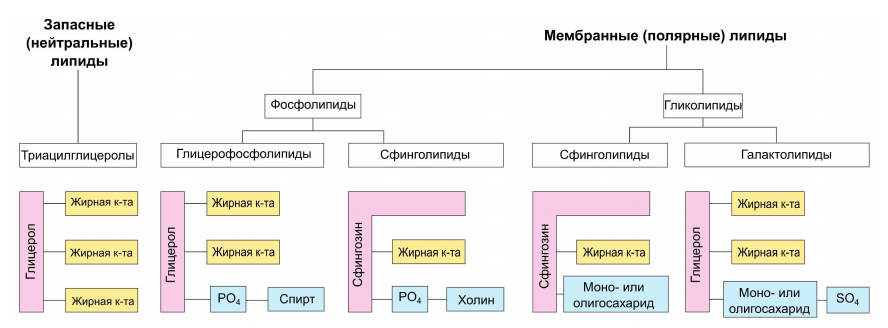
Триацилглицериды – депонируются в жировой ткани. Являются энергетическим запасом организма.
Фосфолипиды – являются структурной основой мембран клеток и мицелл.
Сфингомиелины – основной компонент миелина в нервной ткани, также субстрат для синтеза церамидов и сфингозинов, регулирующих метаболизм.
Гликолипиды (цереброзиды и ганглиозиды) – содержатся в нервной ткани, где отвечают за образование межклеточных контактов.
Переваривание липидов в желудочно-кишечном тракте. Роль желчных кислот и панкреатических липаз: желчные кислоты эмульгируют жиры, а липазы разрушают сложные липиды до более простых.
Значение эмульгирования в переваривании триацилглицеридов: суть в том, что жиры липофильмы и гидрофобны, и потому в водной среде стремятся образовывать крупные капли. Эти крупные капли переваривать можно, но сложно. Желчные кислоты как бы разбивают эти крупные капли на множество мелких, что увеличивает площадь контакта между липидами и ферментами, благодаря чему переваривание проходит хорошо.
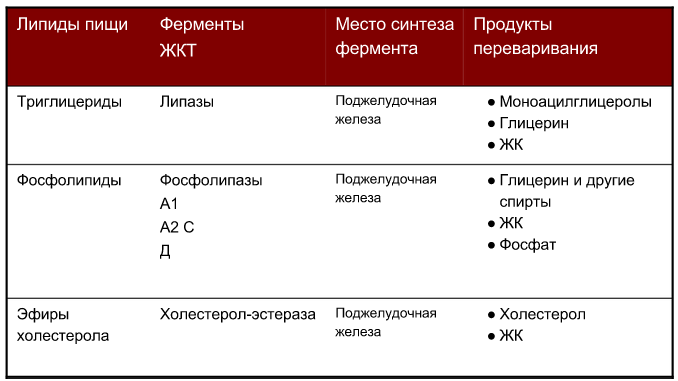
Продукты
переваривания триацилглицеринов,
фосфолипидов, эфиров холестерина:
Нарушения переваривания липидов в желудочно-кишечном тракте: 1. Нарушение секреции желчи. При этом липиды не эмульгируются и поэтому хуже перевариваются и всасываются, из-за чего возникает стеаторея.
2. Нарушение секреции сока поджелудочной железы, что также приводит к стеаторее.
Стеаторея – Повышенной содержание жиров в кале (выше 5 %). При стеаторее нарушается всасывание жирорастворимых витаминов и незаменимых жирных кислот, также нарушается переваривание веществ нелепиднйо природы, поскольку жир обволакивает эти вещества, не давая ферментам расщепить их.
Всасывание продуктов переваривания липидов в тонком кишечнике. Продукты распада жиров (жирные кислоты, 2-моноглицериды, холестерол) при участии солей желчных кислот образуют смешанные мицеллы. Когда мицеллы сближаются с щеточной каёмкой клеток кишечника – липидные компоненты мицелл диффундируют через мембрану. Таким же образом всасываются жирорастворимые витамины и сами соли желчных кислот.
Ресинтез липидов в энтероцитах и его биологическое значение. Всосавшиеся жирные кислоты и 2-моноацилглицерины подвергаются ресинтезу. Вначале ЖК активируются путём связывания с КоА (фермент тиокиназа), затем образовавшийся таким образом ацил-КоА участвует в реакции этерификации 2-моноацилглицерола с образованием вначале диацилглицерола, а потом и триацилглицерола. Реакцию катализирует ацилтрансферазы. Ресинтез нужен нам для того, чтобы всосавшиеся липиды стали «своими».
Гепатоэнтеральная циркуляция желчных кислот. Из холестерина образуются глико(тауро)хенодезоксихолевая и глико(тауро)холевая кислоты, которые первичные желчные кислоты. В кишечнике, под воздействием микрофлоры, первичные желчные кислоты преобразуются во вторичные желчные кислоты (дезоксихолевая и литохолевая кислоты). 95% вторичных желчных кислот всасываются и по воротной вене возращаются в печень, где достраиваются до первичных и снова идут в ход. 5% выводятся с калом.
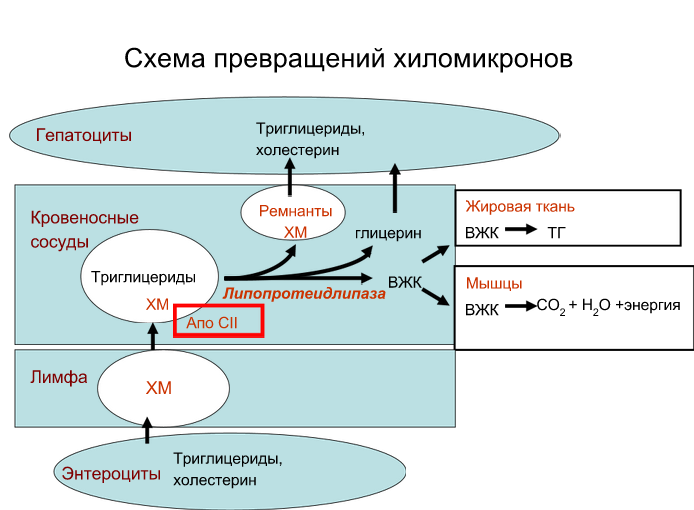
Основные пути превращения хиломикронов.