

kossitsky
.pdfСопоставление частоты разрядов двигательных единиц с частотой, при которой может образоваться гладкий тетанус, позволяет сделать вывод, что в естественных условиях гладкий тетанус может наблюдаться только при очень высокой частоте. Обычным режимом естественного сокращения является зубчатый тетанус или даже ряд последовательных одиночных сокращений двигательной единицы. Тем не менее это не отражается на сокращении целой мышцы; оно, как правило, бывает слитным, напоминающим гладкий тетанус. Причина этого — асинхронность разрядов мотонейронов, а следовательно, и мышечной части двигательных единиц. При отведении игольчатым электродом потенциалов действия одновременно нескольких активных двигательных единиц видна асинхронность их импульсации. В случае электрической активности целой мышцы сложение (интерференция) потенциалов действия многих двигательных единиц дает сложную картину колебаний потенциала, в которой уже не удается различать потенциал каждой из них, а общая частота колебаний существенно превышает частоту разрядов каждой из активных двигательных единиц.
Скорость сокращения двигательных единиц коррелирует и с другими их свойствами. Медленные двигательные единицы, как правило, содержат меньше мышечных волокон и, следовательно, при сокращении развивают меньшую силу. Количество мышечных волокон и развиваемая ими суммарная сила в двигательных единицах одной мышцы могут различаться более чем на порядок. Не менее важно другое различие медленных и быстрых двигательных единиц — устойчивость к утомлению. Медленные двигательные единицы могут работать без утомления гораздо дольше, чем быстрые, что объясняется особенностями их обмена.
Со свойствами мышечных волокон двигательной единицы коррелируют и свойства иннервирующего ее мотонейрона: при естественном напряжении мышцы мотонейроны медленных двигательных единиц обычно оказываются более низкопороговыми, т. е. вовлекаются в возбуждение раньше. Разница в возбудимости мотонейронов позволяет нервной системе дозировать силу сокращения, вовлекая в возбуждения меньшее или большее количество двигательных единиц мышцы. При длительных, но обычно слабых тонических напряжениях, связанных, например, с поддержанием позы, активируются только низкопороговые медленные, устойчивые к утомлению двигательные единицы. Если необходимо осуществить сильное фазное напряжение, в возбуждение вовлекаются высокопороговые, быстрые сильные двигательные единицы.
Рассмотренные двигательные единицы теплокровных животных и человека относятся к классу так называемых фазных двигательных единиц. У амфибий и рептилий, а также в некоторых (немногих) мышцах теплокровных (наружные мышцы глаза) содержатся особые тонические двигательные единицы — мышечные волокна, которые существенно отличаются от волокон фазных единиц. Возбуждение тонических волокон не подчиняется закону «все или ничего» и имеет характер локального ответа, поэтому ограничивается областью нервно-мышечного окончания или тем участком, к которому непосредственно приложено электрическое или химическое раздражение. Охват возбуждением всего волокна возможен потому, что на каждом мышечном волокне имеется не одно, а множество нервных окончаний. Одновременное поступление к этим окончаниям нервного импульса вызывает сокращение всего волокна. Это сокращение существенно медленнее, чем сокращение фазных мышечных волокон.
Регистрация электрической активности двигательных единиц у человека показала, что в естественных условиях мышцы редко бывают полностью расслабленными. Обычно в них наблюдается небольшая, так называемая позная, активность, или позный тонус; при этом низкопороговые медленные двигательные единицы разряжаются с небольшой частотой.
Тонус и особенно его нарушения при ряде заболеваний нервной системы связаны с изменением состояния рефлекторных механизмов, в частности рефлексов с проприорецепторов мышц, повышение возбудимости которых ведет к повышению тонуса.
51

МЕХАНИЗМЫ МЫШЕЧНОГО СОКРАЩЕНИЯ
Структура миофибрилл и ее изменения
при сокращении. Миофибриллы представляют собой сократительный аппарат мышечного волокна. В поперечнополосатых мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими свойствами. Одни участки анизотропны, т. е. обладают двойным лучепреломлением. В обыкновенном свете они выглядят темными, а в поляризованном — прозрачными в продольном направлении и непрозрачными в поперечном. Другие участки в обыкновенном свете выглядят светлыми — они изотропны, т. с. не обладают двойным лучепреломлением (рис. 34, а). Анизотропные участки обозначают буквой А, изотропные — буквой /. В середине диска А различается светлая полоска Н, посередине диска / — темная полоска Z, представляющая собой тонкую мембрану, сквозь поры которой проходят миофибриллы.
Благодаря наличию этой опорной структуры параллельно расположенные однозначные диски отдельных фибрилл
внутри одного волокна во время сокращения не смещаются по отношению друг к другу. Современные представления о структуре миофибриллярного аппарата основываются на исследованиях структуры мышечного волокна при помощи электронной микроскопии, рентгеноструктурного анализа, фазово-контрастной и интерференционной микроскопии в сочетании с гистохимическими методами.
Установлено, что каждая миофибрилла мышечного волокна диаметром около 1 мкм состоит в среднем из 2500 протофибрилл, представляющих собой удлиненные полимеризованные молекулы белков миозина и актина. Миозиновые протофибриллы, или, как их принято обозначать, нити, вдвое толще актиновых. Их диаметр примерно 10 нм. В состоянии покоя мышечного волокна нити расположены в миофибрилле таким образом, что тонкие длинные актиновые нити входят своими концами в промежутки между толстыми и более короткими миозиновыми нитями (рис. 34, б)'. Благодаря этому диски / состоят только из актиновых нитей, а диски А — из нитей миозина, а, возможно, еще и другого белка.
Светлая полоска Н представляет собой узкую зону, свободную от актиновых нитей. Мембрана Z, проходя через середину диска /, скрепляет между собой эти нити. Важным компонентом ультрамикроскопической структуры миофибрилл являются также многочисленные поперечные мостики, соединяющие между собой миозиновые и актиновые нити. При сокращении мышечного волокна указанные нити не укорачиваются, а начинают «скользить» друг по другу: актиновые нити вдвигаются в промежутки между миозиновыми, в результате чего диски / укорачиваются, а диски А сохраняют свой размер. Почти исчезает светлая полоска Н, так как актиновые нити при сокращении сближаются друг с другом своими концами (рис. 34, в). Причиной «скольжения» является химическое взаимодействие между актином и миозином в присутствии ионов Са2+ и АТФ. Наблюдается своего рода химическое «зубчатое колесо», как бы протягивающее одну группу нитей по другой. Роль «зубчиков» в этом процессе приписывают поперечным мостикам, обеспечивающим взаимодействие активных центров белков миозиновых и актиновых нитей.
52
Роль потенциала действия в возникновении мышечного сокращения. В естественных
условиях деятельности скелетной мышцы инициатором ее сокращения является потенциал действия, распространяющийся при возбуждении вдоль поверхностной мембраны мышечного волокна.
Если кончик микроэлектрода при помощи микронанипулнтора приложить к поверхности мышечного волокна лягушки в области диска /, то при нанесении очень слабого электрического стимула, вызывающего деполяризацию, диски / по обе стороны от мембраны Z начнут укорачиваться. При этом, однако, сокращение распространяется не в стороны, а в глубь волокна, вдоль диска /. Приложение слабого стимула к другим участкам миофибриллы подобного эффекта не вызывает. Из этого следует, что деполяризация поверхностной мембраны мышечного волокна в области дисков / является пусковым механизмом сократительного процесса.
Важным промежуточным звеном между деполяризацией мембраны и началом мышечного сокращения является проникновение в область миофибрилл свободных ионов Са2+. В состоянии покоя основная часть ионов Са2+ в скелетном мышечном волокне хранится в так называемом саркоплазматическом ретикулуме. Он представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу.
Вмембране саркоплазматического рётикулума локализованы две важнейшие транспортные
системы, обеспечивающие накопление в ретикулуме ионов Са2'+ («секвестрация» — захват их из миоплазмы) и освобождение Са2+ из ретикулума при возбуждении.
Функцию кальциевого насоса выполняет так называемая Са-зависимая АТФ-аза (Са — АТФ-аза). Энергия, выделяющаяся при
расщеплении АТФ, используется для секвестрации ионов Са2+ в ретикулум. Благодаря этому в
покоящемся волокне концентрация свободных ионов Са2+ в цитоплазме поддерживается на очень низком уровне. Поступая внутрь ретикулума
(главным образом в его продольные трубочки), ионы Са2+ частично связываются белковыми молекулами, устилающими внутреннюю
поверхность его трубочек и цистерн. Концентрация свободных Са2+ в полости ретикулума близка к концентрации их в наружной среде, т. е. во внеклеточной жидкости.
Вмеханизме освобождения ионов Са2+ из
ретикулума при возбуждении особую роль играет система поперечных трубочек (Т-систе-ма),
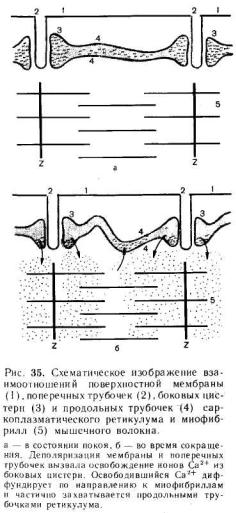
представляющих собой впячивания поверхностной мембраны. Диаметр каждой трубочки около 0,05 мкм. На рис. 35 приведена схема продольного среза через быстрое мышечное волокно лягушки. Видно, что по обе стороны от поперечной трубочки.расположены боковые (терминальные) цистерны ретикулума. Вместе с трубочкой они образуют так называемые триады. Мембрана поперечных трубочек по своим свойствам сходна с поверхностной мембраной; она содержит электровозбудимые натриевые каналы и способна к генерации и проведению потенциала действия. Во время возбуж-
дения потенциал действия с поверхностной мембраны распространяется вдоль мембраны поперечных трубочек в глубь волокна и при помощи особого, пока еще полностью не изученного, механизма вызывает освобождение ионов Са2+ из боковых цистерн. Боковые цистерны расположены таким образом, что освободившиеся ионы Са2+ попадают непосредственно в ту область, где происходит образование актомиозина.
Как отмечалось, начало мышечного сокращения приурочено к первой трети восходящего колена потенциала действия, а именно к моменту, когда внутренний потенциал волокна возрастает с исходных —90 мВ до примерно —50 мВ. Этот потенциал является пороговым для возникновения механического ответа. Предполагают, что именно при достижении указанного уровня деполяризации концентрация свободных ионов Са2+ в миофибрилле достигает критической величины, необходимой для начала взаимодействия актиновых и миозиновых нитей.
Процесс освобождения Са2+ прекращается после окончания пика потенциала действия. Тем не менее сокращение продолжает нарастать до тех пор, пока активация кальциевого насоса ретикулума не вызовет снижения концентрации ионов Са2+ в миоплазме. Тогда сокращение сменяется расслаблением.
Таким образом, последовательность событий, ведущих к сокращению, а затем расслаблению мышечного волокна, представляется в следующем виде: раздражение → возникновение потенциала действия → проведение его вдоль клеточной мембраны и в глубь волокна по трубочкам→освобождение Са2+ из боковых цистерн саркоплазматического ретикулума и диффузия его к миофибриллам→ взаимодействие («скольжение») актиновых и миозиновых нитей, приводящее к укорочению миофибриллы → активация кальциевого насоса→- снижение концентрации свободных ионов Са2+ в саркоплазме → расслабление миофибрилл.
Роль АТФ в механизмах мышечного сокращения
Впроцессе взаимодействия миозиновых и актиновых нитей в присутствии Са2+ важную роль играет богатое энергией соединение — АТФ. Впервые В. А. Энгельгард
иМ. Н. Любимова в 1939 г. нашли, что мышечный белок — миозин — обладает свойствами фермента аденозинтрифосфатазы (АТФ-аза). Эти авторы также обнаружили, что под влиянием АТФ изменяются и механические свойства миозина — резко увеличивается растяжимость его нитей. В последующие годы был открыт белок актин и показано, что он активирует АТФ-азную активность миозина.
Внастоящее время взаимодействие АТФ с сократительными белками и ионами Са2+
представляют следующим образом. В покоящейся мышце, т. е. в условиях, когда концентрация ионов Са2+ в миоплазме очень низкая, взаимодействию миозиновых и актиновых
нитей препятствуют молекулы белка тропонина, расположенные на актиновых нитях. Тропонин обладает очень высоким сродством к ионам Са2+. Как только концентрация Са2+ в миофибриллах повышается, тропонин связывает Са2+ и изменяет свое расположение на актиновой нити таким' образом, что делает возможным ее взаимодействие с миозиновой нитью. Формирующиеся при этом поперечные мостики перемещают актиновую нить лишь на 1 % ее длины. Чтобы обеспечить дальнейшее продвижение нити и соответ-
ственно сокращение волокна, необходимо, чтобы эти мостики разъединились и прикрепились к новым участкам актиновой нити. Такое разъединение мостиков осуществляется при расщеплении молекул АТФ АТФ-азой миозина. Таким образом, взаимодействие миозина и актина, возможное при связывании Са2+ тропонином, активирует АТФ-азу миозина, последняя расщепляет АТФ, а это приводит к разъединению миозина и актина. В присутствии Са2+ и АТФ в миоплазме этот процесс многократно повторяется: мостики повторно образуются и расходятся, в результате чего актиновые нити «скользят» и мышечное волокно укорачивается.
Подсчитано, что при изотоническом сокращении скелетной мышцы лягушки поперечные мостики должны совершить за 0,1 с 50 таких движений, чтобы длина каждого
54
саркомера волокна укоротилась на 50 %. При каждом движении мостиков происходит расщепление молекул АТФ.
Таким образом, присутствие АТФ в мышце является обязательным условием для обратимости связывания актина и миозина. Трупное окоченение возникает при условии, если концентрация АТФ в мышце падает ниже некоторой критической величины. Тогда комплекс актин — миозин становится стабильным.
Итак, энергия АТФ используется во время деятельности скелетной мышцы для трех основных процессов: 1) работы натр ий-калиевого насоса, обеспечивающего поддержание постоянства градиента концентрации ионов Na+ и К+ по обе стороны мембраны; 2) про-
цесса «скольжения» актиновых и миозиновых нитей, ведущего к укорочению миофиб-
рилл; 3) работы кальциевого насоса, необходимого для расслабления волокна. В соответствии с этим ферменты, расщепляющие АТФ, локализованы в трех различных структурах мышечного волокна: клеточной мембране, миозиновых нитях и мембранах саркоплазматического ретикулума.
Ресинтез АТФ, непрерывно расщепляющейся в процессе деятельности мышцы, осуществляется двумя основными путями. Первый состоит в ферментативном переносе фосфатной группы от богатого энергией фосфорного соединения креатинфосфата на аденозиндифосфорную кислоту. Креатинфосфат содержится в мышце в значительно больших количествах, чем АТФ, и обеспечивает ее ресинтез в течение тысячных долей секунды. Однако при интенсивной работе мышцы запасы креатинфосфата быстро истощаются, поэтому важен второй путь—более медленный ресинтез АТФ. Он связан с гликолитическими и окислительными процессами, протекающими в мышце как в условиях покоя, так и особенно интенсивно во время деятельности. Окисление молочной и пировиноградной кислот, образующихся в мышце во время ее сокращения, сопровождается фосфорилированием аденозиндифосфорной кислоты и креатина, т. е. ресинтезом креатинфосфата и АТФ.
Нарушение ресинтеза АТФ ядами, подавляющими гликолитические и окислительные процессы, ведет к полному исчезновению АТФ и креатинфосфата, вследствие чего кальциевый насос перестает работать. Концентрация Са2+ в области миофибрилл значительно возрастает и мышца приходит в состояние длительного необратимого укорочения.
Теплообразование при сократительном процессе и энергия сокращения
Образование тепла в мышечной ткани при работе было открыто Гельмгольцем и В. Я. Данилевским во второй половине XIX в. В дальнейшем Хиллу с сотр. удалось создать высокочувствительные приборы, которые позволили зарегистрировать и измерить теплопродукцию мышц и нервов в покое и при возбуждении.
По своему происхождению и времени развития теплообразование было подразделено Хиллом на две основные фазы. Первая фаза приблизительно в 1000 раз короче второй и называется фазой начального теплообразования. Она начинается с момента возбуждения мышцы и продолжается в течение всего сокращения, включая фазу расслабления.
В свою очередь начальное теплообразование может быть разделено на несколько частей: а) тепло активации; б) тепло укорочения; в) тепло расслабления.
Тепло активации освобождается непосредственно после нанесения раздражения, но до сколько-нибудь различимого сокращения мышечных волокон. Поэтому указанная порция тепла рассматривается как тепловой эффект тех химических процессов, которые переводят мышцу из невозбужденного состояния в активное. При тетаническом сокращении тепло активации выделяется в течение всего времени раздражения мышцы при каждом потенциале действия.
Тепло укорочения обусловлено самим сократительным процессом. Если путем сильного растяжения мышцы воспрепятствовать ее сокращению, эта порция тепла не выделяется.
55
Тепло расслабления связано с освобождением энергии в результате расслабления мышцы. Если мышца подняла груз во время сокращения, то по окончании его количество выделяемого тепла увеличивается.
Вторая фаза теплопродукции длится несколько минут после расслабления и носит название запаздывающего, или восстановительного, теплообразования.
Она связана с химическими процессами, обеспечивающими ресинтез АТФ. В опытах на мышцах, сокращающихся в отсутствие кислорода, Хилл показал, что в отличие от начального теплообразования, для которого кислород не нужен, 90 % восстановительного тепла образуется в результате окислительных процессов и лишь 10 % этого тепла обусловлены анаэробными процессами обмена веществ. Тепло восстановления по своей величине примерно равно количеству тепла, выделяемого мышцей во время сокращения. Это соответствие становится понятным, если учесть, что химические процессы, обусловливающие восстановительное теплообразование, направлены на ресинтез АТФ, являющийся основным непосредственным источником энергии мышечного сокращения. Главную роль в ресинтезе АТФ и восстановительном теплообразовании играют процессы гликолиза и окислительного фосфорилирования. Отравление мышцы монойодуксусной кислотой, прекращающей гликолитическое образование молочной и пировиноградной кислот, почти полностью выключает запаздывающее теплообразование и ресинтез АТФ даже в присутствии кислорода.
РАБОТА И СИЛА МЫШЦ
Величина сокращения (степень укорочения) мышцы при данной силе раздражения (т. е. при данном числе активированных волокон) зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект; при сильном растяжении сокращение мышцы ослабляется. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения снижается.
Для измерения силы мышцы определяют либо тот максимальный груз, который она
всостоянии поднять, либо максимальное напряжение, которое она может развить в условиях изометрического сокращения. Эта сила может быть очень велика. Установлено, что собака мышцами челюсти может поднять груз, превышающий массу ее тела в 8,3 раза.
Одиночное мышечное волокно способно развить напряжение 100—200 мг. Учитывая, что общее количество мышечных волокон в теле человека приблизительно 15—30 млн., они могли бы развить напряжение в 20—30 т, если бы все одновременно тянули в одну сторону.
Сила мышцы при прочих равных условиях зависит от ее поперечного сечения. Чем больше физиологическое поперечное сечение мышцы, т. е. сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косым расположением волокон сумма поперечных сечений может значительно превышать геометрическое поперечное сечение самой мышцы (рис. 36). По этой причине сила мышцы с косо расположенными волокнами значительно больше, чем сила мышцы той же толщины, но при продольном расположении волокон. Чтобы иметь возможность сравнивать силу разных мышц, максимальный груз, который
всостоянии поднять мышца, делят на площадь ее физиологического поперечного сечения. Таким образом вычисляют абсолютную мышечную силу. Абсолютная сила, выраженная
вкилограммах на I см2, икроножной мышцы человека равна 5,9, сгибателя плеча — 8,1, жевательной мышцы — 10, двуглавой мышцы плеча — 11,4, трехглавой мышцы плеча — 16,8, гладких мышц— 1.
Примером мышц с продольным расположением волокон может служить m.sartorius, с косым — m.m.intercostales. Большинство мышц млекопитающих и человека перистого
56

строения. Перистая мышца имеет большое физиологическое сечение, а потому обладает большой силой.
Зависимость изометрического напряженияот исходной длины мышцы
Если покоящуюся мышцу постепенно растягивать за сухожильные концы, то в силу эластических свойств напряжение мышцы будет возрастать так, как это показано на рис. 37 (кривая А).
Нанесение электрических раздражений на мышцу вызывает прирост напряжения (кривая Б), величина которого с увеличением исходной длины мышечных волокон вначале возрастает, а затем уменьшается. Если сопоставить кривую Б с кривой А, то можно получить представление о зависимо-
сти этого активного прироста изометрического напряжения от длины мышечных саркомеров. При некоторой средней длине саркомеров сила, развиваемая мышечным волокном, максимальна. При перерастяжении волокон эта сила уменьшается. Причина такой зависимости напряжения от исходной длины саркомеров становится ясной при рассмотрении взаимного расположения миозиновых и актиновых нитей при разной длине саркомеров (рис. 37,В).
Работа мышцы измеряется произведением поднятого груза на величину укорочения мышцы. Между грузом, который поднимает мышца, и выполняемой ею работой существует следующая зависимость. Внешняя работа мышцы равна нулю, если мышца сокращается без нагрузки. По мере увеличения груза работа сначала увеличивается, а затем
57

постепенно уменьшается. При очень большом грузе, который мышца неспособна поднять, работа становится равной нулю. На рис. 38 показаны соотношения между величиной нагрузки, степенью укорочения мышцы и величиной работы. Наибольшую работу мышца совершает при некоторых средних нагрузках, в данном случае при 200—250 г. Мощность мышцы, измеряемая величиной работы в единицу времени, также достигает максимальной величины при средних нагрузках. Поэтому зависимость работы и мощности от нагрузки получила название правила средних нагрузок.
Работа мышцы, при которой происходит перемещение груза и движение костей в суставах, называется динамической. Работа мышцы, при которой мышечные волокна развивают напряжение, но почти не укорачиваются (это происходит, когда мышца сокращается в изометрическом режиме), называется статической. Примером является работа по удержанию груза. Статическая работа более утомительна, чем динамическая. Работа может совершаться и в условиях удлинения мышцы (опускание груза), тогда она называется уступающей работой.
Утомление мышцы
Утомлением называется временное понижение работоспособности клетки, органа или целого организма, наступающее в результате работы и исчезающее после отдыха.
Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, к которой подвешен небольшой груз, то амплитуда ее сокращений постепенно убывает до нуля. Регистрируемую при этом запись сокращений называют кривой утомления.
Наряду с изменением амплитуды сокращений при утомлении нарастает латентный период сокращения и удлиняется период расслабления мышцы. Однако все эти изменения возникают не тотчас после начала работы, а спустя некоторое время, в течение которого наблюдается увеличение амплитуды одиночных сокращений мышцы. Этот период называется периодом врабатывания. При дальнейшем длительном раздражении развивается утомление мышечных волокон.
Понижение работоспособности изолированной мышцы при ее длительном раздражении обусловлено двумя основными причинами. Первой из них является то, что во время сокращения в мышце накапливаются продукты обмена веществ (фосфорная, молочная кислоты и др.), оказывающие угнетающее действие на работоспособность мышечных волокон. Часть этих продуктов, а также ионы калия диффундируют из волокон наружу в околоклеточное пространство и оказывает угнетающее влияние на способность возбудимой мембраны генерировать потенциалы действия. Если изолированную мышцу, помещенную в небольшой объем жидкости Рингера, длительно раздражая довести до полного утомления, то достаточно только сменить омывающий ее раствор, чтобы восстановились сокращения мышцы.
Другая причина развития утомления изолированной мышцы — постепенное исто-
58

щение в ней энергетических запасов. При длительной работе изолированной мышцы происходит резкое уменьшение запасов гликогена, вследствие чего нарушаются процессы ресинтеза АТФ и креатинфосфата, необходимых для осуществления сокращения.
Следует подчеркнуть, что утомление изолированной скелетной мышцы при ее прямом раздражении является лабораторным феноменом. В естественных условиях утомление двигательного аппарата при длительной работе развивается более сложно и зависит от большого числа факторов. Обусловлено это, во-первых, тем, что в организме
мышца непрерывно снабжается кровью и, следовательно, получает с ней определенное количество питательных веществ (глюкоза, аминокислоты) и освобождается от продуктов обмена, нарушающих нормальную жизнедеятельность мышечных волокон. Вовторых, в целом организме утомление зависит не только от процессов в мышце, но и от процессов, развивающихся в нервной системе, участвующих в управлении двигательной деятельностью. Так, например, утомление сопровождается дискоординацией движений, возбуждением многих мышц, которые не участвуют в совершении работы.
И. М. Сеченов (1903) показал, что восстановление работоспособности утомленных мышц руки человека после длительной работы по подъему груза ускоряется, если в период отдыха производить работу другой рукой. Временное восстановление работоспособности мышц утомленной руки может быть достигнуто и при других видах двигательной активности, например при работе мышц нижних конечностей. В отличие от простого покоя такой отдых был назван И. М. Сеченовым активным. Он рассматривал эти факты как доказательство того, что утомление развивается прежде всего в нервных центрах.
Убедительным доказательством роли нервных центров в развитии утомления могут служить опыты с внушением. Так, находясь в состоянии гипноза, испытуемый может длительное время поднимать тяжелую гирю, если ему внушить, что в его руке находится легкая корзина. Наоборот, при внушении испытуемому, что ему дана тяжелая гиря, утомление быстро развивается при подъеме легкой корзины. При этом изменения пульса, дыхания и газообмена находятся в соответствии не с реальной работой, осуществляемой человеком, а с той, которая ему внушена.
При выявлении причин утомления двигательного аппарата в применении к целостному организму в настоящее время нередко различают два вида двигательной деятельности: локальную, когда активно сравнительно небольшое количество мышц, и общую, когда большинство мышц тела участвует в работе. В первом случае среди причин утомления на первое место выступают периферические факторы, т. е. процессы в самой мышце; во втором ведущее значение приобретают центральные факторы и недостаточность. вегетативного обеспечения движений (дыхания, кровообращения). Исследованию механизмов утомления большое внимание уделяется в физиологии труда и спорта.
Эргография. Для изучения мышечного утомления у человека в лабораторных условиях пользуются эргографами — приборами для записи механограммы при движениях, ритмически выполняемых группой мышц. Такая запись позволяет определить количество выполняемой работы.
Примером такого простейшего прибора может служить эргограф Моссо, записывающий движение нагруженного пальца. Сгибая и разгибая палец при фиксированном положении руки, испытуемый поднимает и опускает подвешенный к пальцу груз в определенном, заданном ритме (например, в ритме ударов метронома).
Существуют эргографы, воспроизводящие те или иные рабочие движения человека. Так, широко используются велоэргографы (велоэргометры). Человек ногами вращает педали прибора при различном, заданном заранее сопротивлении этому движению. Специальные датчики позволяют регистрировать параметры движения и количество выполненной работы. Одновременно можно
59
регистрировать показатели дыхания, кровообращения, ЭКГ. Велоэргографы широко используются в медицине для определения функциональных возможностей организма человека.
Форма эргограммы и величина работы, произведенной человеком до наступления утомления, варьируют у разных лиц и даже у одного и того же лица при различных условиях. В этом отношении показательны эргограммы, записанные Моссо на самом себе до и после приема зачета у студентов. Эти эргограммы свидетельствуют о резком уменьшении работоспособности после напряженной умственной работы (рис. 39).
Рабочая гипертрофия мышц и атрофия от бездеятельности
Систематическая интенсивная работа мышцы способствует увеличению массы мышечной ткани. Это явление названо рабочей гипертрофией мышцы. В основе гипертрофии лежит увеличение массы цитоплазмы мышечных волокон и числа содержащихся
вних миофибрилл, что приводит к увеличению диаметра каждого волокна. При этом
вмышце происходит активация синтеза нуклеиновых кислот и белков и повышается содержание веществ, доставляющих энергию, используемую при мышечном сокращении,— аденозинтрифосфата и креатинфосфата, а также гликогена. В результате сила и скорость сокращения гипертрофированной мышцы возрастают.
Увеличению числа миофибрилл при гипертрофии способствует преимущественно статическая работа, требующая большого напряжения (силовая нагрузка). Даже кратковременных упражнений, проводимых ежедневно в условиях изометрического
режима, достаточно для того, чтобы увеличилось количество миофибрилл. Динамическая мышечная работа, производимая без особых усилий, не вызывает гипертрофии мышцы.
У тренированных людей, у которых многие мышцы гипертрофированы, мускулатура может составлять до 50 % массы тела (вместо 35—40 % в норме).
Противоположным рабочей гипертрофии состоянием является атрофия мышц от бездеятельности. Она развивается во всех случаях, когда мышца почему-либо длительно не совершает нормальной работы. Это наблюдается, например, при обездвижении конечности в гипсовой повязке, долгом пребывании больного в постели, перерезке сухожилия, вследствие чего мышца перестает совершать работу, и т. п.
При атрофии диаметр мышечных волокон и содержание в них сократительных белков, гликогена, АТФ и других важных для сократительной деятельности веществ уменьшаются. После возобновления нормальной работы мышцы атрофия постепенно исчезает.
Особый вид мышечной атрофии наблюдается при денервапии мышцы, т. е. после утраты ее связи с нервной системой, например при перерезке се двигательного нерва. Этот вид атрофии рассмотрен далее.
ГЛАДКИЕ МЫШЦЫ
Функции гладких мышц в разных органах
Гладкие мышцы в организме высших животных и человека находятся во внутренних органах, сосудах и коже. Они способны осуществлять относительно медленные движения и длительные тонические сокращения.
Относительно медленные, часто имеющие ритмический характер сокращения гладких мышц стенок полых органов: желудка, кишок, протоков пищеварительных желез, мочевого пузыря, желчного пузыря и др., обеспечивают перемещение содержимого этих органов. Примером являются маятникообразные и перистальтические движения кишечника.
Длительные тонические сокращения гладких мышц особенно резко выражены в сфинктерах полых органов; их тоническое сокращение препятствует выходу содержимого. Этим обеспечивается накопление желчи в желчном пузыре и мочи в мочевом пузыре, оформление каловых масс в толстой кишке и т. д.
60