

kossitsky
.pdfи любое раздражение кожи начинает вызывать вместо обычного рефлекса усиление тонического обнимательного рефлекса. После кастрации эти рефлексы угасают, но вновь восстанавливаются при введении полового гормона тестостерона. Показательно, что усиление активности нейронов моторных центров развивается даже при изоляции спинного мозга и добавлении тестостерона в перфузирующий раствор.
Взаимодействие рефлексов может проявляться как во взаимном облегчении (суммация), так и угнетении (окклюзия). Последнее явление выражается в том, что суммарный результат оказывается значительно меньшим, чем сумма взаимодействующих реакций. Согласно Ч. Шерриигтону (рис. 75), явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными частями взаимодействующих рефлексов. Поэтому при одновременном поступлении двух афферентных влияний ВПСП вызывается каждым из них отчасти в одних и тех же мотонейронах.
ОБЪЕДИНЕНИЕ НЕЙРОНОВ В НЕРВНЫЙ ЦЕНТР
В результате интегративной деятельности нейронов ЦНС, в основе которой лежат процессы суммации возбуждающих и тормозных постсинаптических потенциалов, возникающих в разных участках поверхностной мембраны, в нервной клетке происходит генерация импульсов или команд, регулирующих деятельность других нейронов или органов. В этом смысле работа индивидуального нейрона в какой-то степени подобна работе нервной системы в целом. Однако сложность и многообразие задач, выполняемых нервной системой, приводит к высокой специализации отдельных нейронов, и не только отдельных нейронов, но и нейронных объединений — нервных центров, деятельность которых связана с осуществлением различных функций. Функционирование ЦНС в целом осуществляется благодаря деятельности значительного числа таких центров, представляющих собой ансамбли нервных клеток, объединенных с помощью синаптических контактов и отличающихся огромным разнообразием и сложностью внутренних и внешних связей.
В соответствии с выполняемой функцией можно выделить различные чувствительные центры, центры вегетативных функций, двигательные центры, центры психических функций и т. д. Различные нервные центры характеризуются определенной топографией своего расположения в пределах ЦНС и более подробно будут рассмотрены в следующей главе.
Глава 6
ЧАСТНАЯ ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
В основе современных представлений о структуре и функции мозга лежит нейронная теория, рассматривающая ЦНС как совокупность синаптически связанных между собой нервных элементов — нейронов. Однако, несмотря на то что нервные клетки, из которых построен мозг, составляют единую особым образом организованную сеть, сложность ЦНС настолько велика, что для анализа ее функциональных свойств целесообразно разделение всего мозга на определенные области или отделы в соответствии с их макроструктурой, развитием, функцией и нейронной организацией. Такое подразделение в известной мере условно, так как нейроны, находящиеся в отдаленных друг от друга отделах мозга (например, гигантские пирамидные клетки двигательной коры и мотонейроны спинного мозга), бывают связаны между собой значительно более тесно, чем нейроны территориально близко расположенные, в пределах одной и той же области, но отно-
111
сящиеся к различным физиологическим системам. Несмотря на это, разделение ЦНС на определенные отделы удобно как для нейрофизиологического анализа, так и для клинических целей.
Выяснение специфических особенностей различных нервных образований и закономерностей их взаимодействия относится к частной физиологии ЦНС.
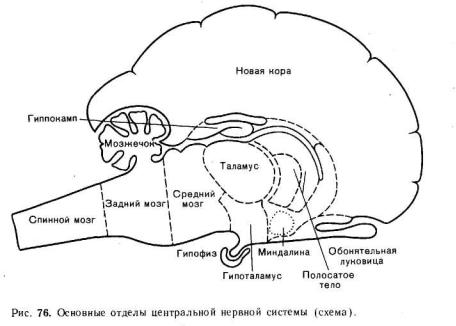
Схематическое изображение ЦНС и входящих в ее состав отделов показано на рис. 76. На этой схеме в общем виде обозначены спинной мозг, задний мозг (включающий продолговатый мозг и варолиев мост или мост мозга), средний мозг, мозжечок, промежуточный мозг (таламус и гипоталамус) и передний мозг (включающий стриопаллидарную систему подкорковых ядер и кору больших полушарий).
СПИННОЙ МОЗГ
Спинной мозг — наиболее древний отдел ЦНС позвоночных, появившийся уже у наиболее примитивных представителей хордовых — ланцетника — еще до сформирования головного мозга. Поэтому он более, чем другие отделы ЦНС, сохранил черты древней организации, выражающиеся в метамерном строении.
Спинной мозг является самым каудальным отделом ЦНС. Он помещается в позвоночном канале и имеет неодинаковую длину у разных представителей позвоночных. У большинства рыб, хвостатых амфибий, рептилий и птиц он заполняет весь позвоночный канал, в то время как у бесхвостых амфибий и млекопитающих — только его часть. Поэтому в каудальном отделе позвоночного канала собираются корешки каудальных сегментов спинного мозга, образуя так называемый конский хвост.
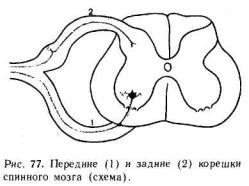
Спинной мозг характеризуется выраженным сегментарным строением, отражающим сегментарное строение тела позвоночных. От каждого спинномозгового сегмента отходят две пары вентральных и дорсальных корешков (рис. 77). Дорсальные корешки формируют афферентные входы спинного мозга. Они образованы центральными отростками волокон первичных афферентных нейронов, тела которых вынесены на периферию и находятся в спинномозговых ганглиях. Вентральные корешки образуют эфферентные выходы спинного мозга. В них проходят аксоны α- и γ-мотонейронов, а также преганглио-
112
нарных нейронов вегетативной нервной системы. Такое распределение афферентных и эфферентных волокон было установлено еще в начале прошлого века и получило название
закона Белла — Мажанди. После перерезки передних корешков на одной стороне наблюдается полное выключение двигательных реакций; но чувствительность этой стороны тела сохраняется. Перерезка задних корешков выключает чувствительность, но не приводит к утрате двигательных реакций мускулатуры.
Нейронная организация. Нейроны спин-
номозговых ганглиев относятся к простым униполярным, или псевдоуниполярным, нейронам. Название «псевдоуниполярный»
объясняется тем, что в эмбриональном периоде первичные афферентные нейроны происходят от биполярных клеток, отростки которых затем сливаются. Нейроны спинномозговых ганглиев можно подразделить на клетки малых и больших размеров. Тело крупных нейронов имеет диаметр порядка 60—120 мкм, в то время как у мелких нейронов он колеблется от 14 до 30 мкм.
Крупные нейроны дают начало толстым миелинизированным волокнам. От мелких начинаются как тонкие миелинизированные, так и немиелинизированные волокна. После бифуркации оба отростка направляются в противоположные направления: центральный входит в дорсальный корешок и в его составе — в спинной мозг, периферический — в различные соматические и висцеральные нервы, подходящие к рецепторным образованиям кожи, мышц и внутренних органов.
Иногда центральные отростки первичных афферентных нейронов заходят в вентральный корешок. Это происходит при трифуркации аксона первичного афферентного нейрона, в результате которой его отростки проецируются в спинной мозг и через дорсальный и вентральный корешки.
Из всей популяции клеток дорсальных ганглиев примерно 60—70% относится к мелким нейронам. Это соответствует тому, что число немиелинизированных волокон в дорсальном корешке превышает число миелинизированных волокон.
Тела нейронов спинномозговых ганглиев не имеют дендритных отростков и не получают синаптических входов. Их возбуждение происходит в результате прихода потенциала действия по периферическому отростку, контактирующему с рецепторами.
Клетки спинномозговых ганглиев содержат высокие концентрации глутаминовой кислоты — одного из предполагаемых медиаторов. Их поверхностная мембрана содержит рецепторы, специфически чувствительные к γ-аминомасляной кислоте, что совпадает с высокой чувствительностью к 7-аминомасляной кислоте центральных окончаний первичных афферентных волокон. Малые нейроны ганглиев содержат вещество Р или соматостатин. Оба этих полипептида также являются вероятными медиаторами, высвобождаемыми окончаниями первичных афферентных волокон.
Каждая пара корешков соответствует одному из позвонков и покидает позвоночный канал через отверстие между ними. Поэтому сегменты спинного мозга принято обозначать по тому позвонку, возле которого из спинного мозга выходят соответствующие корешки. Спинной мозг принято также разделять на несколько отделов: шейный, грудной, поясничный и крестцовый, каждый из которых содержит по нескольку сегментов. В связи с развитием конечностей нейронный аппарат тех сегментов спинного мозга, которые их иннервируют, получил наибольшее развитие. Это нашло свое отражение в образовании шейного и поясничного утолщений. В области утолщений спинного мозга корешки содержат наибольшее количество волокон и имеют наибольшую толщину.
На поперечном срезе спинного мозга ясно выделяется центрально расположенное серое вещество, образованное скоплением нервных клеток, и окаймляющее его белое вещество, образованное нервными волокнами. В сером веществе различают вентральные и дорсальные рога, между которыми лежит промежуточная зона. Кроме того, в грудных сегментах различают также боковое выпячивание серого вещества — боковые рога.
113
Все нейронные элементы спинного мозга могут быть подразделены на 4 основные группы:
эфферентные нейроны, вставочные нейроны, нейроны восходящих трактов и интраспи-нальные волокна чувствительных афферентных нейронов.
Моторные нейроны сосредоточены в передних рогах, где они образуют специфические ядра, все клетки которых посылают свои аксоны к определенной мышце. Каждое двигательное ядро обычно тянется на несколько сегментов. Поэтому и аксоны мотонейронов, ин-нервирующих одну и ту же мышцу, покидают спинной мозг в составе нескольких вентральных корешков.
Кроме моторных ядер, расположенных в вентральных рогах, выделяются большие скопления нервных клеток, в промежуточной зоне серого вещества. Это основное ядро вставочных нейронов спинного мозга. Аксоны вставочных нейронов распространяются как внутри сегмента, так и в ближайшие соседние сегменты.
Характерное скопление нервных клеток занимает также дорсальную часть дорсального рога. Эти клетки образуют густые переплетения,
а указанная зона получила название желатинозной субстанции Роланда.
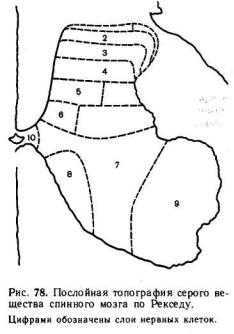
Наиболее точное и систематизированное представление о топографии нервных клеток серого вещества спинного мозга дает разделение его на последовательные слои, или пластины, в каждой из которых группируются главным образом однотипные нейроны.
Хотя послойная топография серого вещества была первоначально выявлена в спинном мозге кошки, она оказалась достаточно универсальной и вполне применима к спинному мозгу как других позвоночных, так и человека.
Согласно этим данным, все серое вещество можно разделить на 10 пластин (рис. 78). Самая первая дорсальная пластина содержит главным образом так называемые краевые нейроны. Их аксоны проецируются рострально, давая начало спиноталамическому тракту. На краевых нейронах оканчиваются волокна тракта Л иссауэра, который образован смесью первичных афферентных волокон и аксонами проприоспинальных нейронов.
Вторая и третья пластины образуют желатинозную субстанцию. Здесь локализуются два основных типа нейронов: более мелкие и относительно крупные нейроны. Хотя тела нейронов второй пластины имеют небольшой диаметр, их дендритные разветвления весьма многочисленны. Аксоны нейронов второй пластины проецируются на тракт Лиссауэра и собственный дорсолатеральный пучок спинного мозга, но многие остаются в пределах желатинозной субстанции. На клетках второй и третьей пластин оканчиваются волокна первичных афферентных нейронов, преимущественно кожной и болевой чувствительности.
Четвертая пластина занимает примерно центр дорсального рога. Дендриты нейронов IV слоя проникают в желатинозную субстанцию, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции и первичных афферентных нейронов.
В целом нервные клетки первой — четвертой пластин захватывают всю вершину дорсального рога и образуют первичную сенсорную область спинного мозга. Сюда проецируются волокна большей части дорсально-корешковых афферентов от экстерорецепторов, включая кожную и болевую чувствительность. В этой же зоне локализованы нервные клетки, дающие начало нескольким восходящим трактам.
114
Впятой и шестой пластинах локализуются многочисленные типы вставочных нейронов, получающие синаптические входы от волокон заднего корешка и нисходящих путей, в особенности кортико-спинального и руброспинального тракта.
Вседьмой и восьмой пластинах локализуются проприоспинальные вставочные нейроны, дающие начало длинным аксонам, достигающим нейронов отдаленных сегментов. Здесь заканчиваются афферентные волокна от проприорецепторов, волокна вестибулоспинального и ретикулоспинального трактов, аксоны проприоспинальных нейронов.
Вдевятой пластине располагаются тела α- и γ-мотонейронов. Этой области достигают также пресинаптические окончания первичных афферентных волокон от мышечных рецепторов растяжения, окончания волокон нисходящих трактов, кортико-спинальных волокон, терминали аксонов возбуждающих и тормозящих вставочных нейронов.
Десятая пластина окружает спинномозговой канал и содержит наряду с нейронами значительное количество глиальных клеток и комиссуральных волокон.
СВОЙСТВА НЕРВНЫХ ЭЛЕМЕНТОВ СПИННОГО МОЗГА
Мотонейроны, расположенные в девятой пластине серого вещества, подразделяются на два основных типа: а- и умотонейроны. Альфа-мотонейроны иннервируют волокна скелетной мускулатуры (экстрафузальные волокна), обеспечивая мышечные сокращения. Гамма-мотонейроны иннервируют рецепторы растяжения (интрафузальные волокна). Благодаря сочетанной активации а- и умотонейронов рецепторы растяжения могут активироваться не только во время растяжения мышц, но и при их сокращении, что важно для обеспечения моторной координации.
Обнаружен также тип мотонейронов — р-мотонейроны, концевые разветвления которых иннервируют параллельно экстрафузальные и интрафузальные волокна. Однако свойства этих мотонейроновизучены еще недостаточно.
Альфа-мотонейроны представляют собой крупные клетки с длинными дендритами. На соме и дендритах α-мотонейронов размещается до 10 000—20 000 синапсов, большая часть которых образована окончаниями интраспинальных вставочных нейронов. Часть синапсов на α-мотонейронах представляет собой контакты с окончаниями первичных афферентных волокон, идущих от мышечных рецепторов растяжения, и с волокнами длинных нисходящих трактов: кортико-спинального, руброспинального, вестибулоспинального и ретикулоспинального.
Аксоны α-мотонейронов характеризуются высокой скоростью проведения — 70— 120 м/с, что согласуется с их значительным диаметром. Имеется четкая корреляция между размерами сомы мотонейронов, толщиной их миелинизированных аксонов и скоростью проведения по аксонам.
Деполяризация, возникающая в α-мотонейронах при активации возбуждающих синаптических входов, вызывает ритмические разряды потенциалов действия. Частота импульсации пропорциональна степени деполяризации, однако обычно она не превышает 10—20 имп/с, что обусловлено интенсивной следовой гиперполяризацией, развивающейся после каждого потенциала действия.
Как и α-мотонейроны, γ мотонейроны расположены в девятой пластине, где они рассеяны среди α-мотонейронов. Гамма-мотонейроны представляют собой значительно более мелкие клетки. Диаметр их тела не превышает 30—40 мкм. В отличие от α-мотоней- ронов γ-мотонейроны не имеют непосредственного синаптического контакта с первичными афферентными волокнами. Однако они моносинаптически активируются волокнами нисходящих трактов, что играет важную роль в α-, γ сопряжении (сочетанной активации).
В соответствии с незначительными размерами тела, аксоны γ мотонейронов также имеют небольшой диаметр. Поэтому скорость проведения по γ волокнам составляет всего 10—40 м/с. Следовая гиперполяризация в γ-мотонейронах выражена незначительно и не ограничивает существенно частоты их импульсации. В этой связи γ мртонейроны могут разряжаться с частотой 300—500 имп/с.
115
Особую группу эфферентных нейронов спинного мозга составляют преганглионар-
ные нейроны вегетативной нервной системы, расположенные как в боковых, так и в перед-
них рогах серого вещества. Аксоны данных нейронов направляются к клеткам ганглиев симпатической цепочки и к интрамуральным ганглиям внутренних органов.
Вставочные, или промежуточные, нейроны спинного мозга представляют довольно разнородную группу нервных клеток, тела, дендриты, аксоны которых находятся впределах.спинного мозга. Вставочные нейроны имеют синаптические контакты только с другими нейронами и составляют значительное большинство нервных элементов спинного мозга.
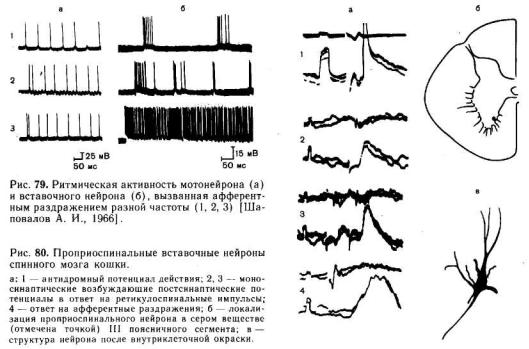
Несмотря на то что различные вставочные нейроны значительно отличаются друг от друга размерами, характером дендритных разветвлений, аксональными проекциями и функциональными особенностями, им присущи и некоторые общие свойства. Как правило, вставочные нейроны имеют тело меньшего диаметра, чем α-мотонейроны. . Следовая гиперполяризация в большинстве разновидностей вставочных нейронов выражена незначительно. Поэтому они способны к генерации высокочастотной импульсной активности. Как видно на рис. 79, частота импульсации вставочного нейрона значительно превышает частоту импульсации мотонейронов. Даже в ответ на одиночный стимул они обычно генерируют множественные разряды. Частота импульсов, регистрируемых в теле некоторых вставочных нейронов, может превышать 1000 в секунду.
К числу промежуточных нейронов относятся описанные выше клетки Реншоу (тормозные вставочные нейроны), получающие возбуждение от афферентных волокон мышечных рецепторов (проприоспинальные вставочные нейроны).
Тормозные вставочные нейроны включены в путь реципрокного торможения мотонейронов мышц-антагонистов.
Для пройриоспинальных вставочных нейронов характерно свойство посылать длинные аксоны, проходящие через несколько сегментов и оканчивающиеся на мотонейронах. Вместе с тем на проприоспинальных клетках конвергируют волокна различных нисходящих трактов. Поэтому они являются важной релейной станцией на пути от вышележащих центров к мотонейронам. Рис. 80 демонстрирует пример моносинаптических ответов,
116
возникающих в проприоспинальном нейроне при раздражении ретикулярной формации. Здесь же показана структура и местоположение этого нейрона.
Нейроны восходящих трактов также целиком находятся в пределах ЦНС. Тела этих клеток расположены в сером веществе спинного мозга, в то время как их аксоны проецируются к ядрам ростральных сегментов спинного мозга и к нейронам различных вышележащих образований. Наиболее хорошо изучены свойства нейронов дорсального и вентрального спинно-мозжечкового трактов, первые из которых, образуя так называемое грудное ядро (ядро Кларка), получают мощное возбуждающее влияние через афферентные пути мышечных рецепторов.
Наконец, центральные окончания первичных афферентных волокон характеризуются следующими особенностями. После вступления в спинной мозг афферентное волокно, как правило, дает начало восходящей и нисходящей ветвям, которые могут идти на значительные расстояния по спинному мозгу. Достигнув уровней конечного ветвления, коллатерали первичных афферентных волокон проникают в серое вещество и образуют концевые ветвления, которые заканчиваются синаптическими образованиями на телах и дендритах иннервируемых ими клеток. Концевые разветвления одного первичного афферентного волокна имеют многочисленные синапсы на одном мотонейроне. Кроме того, каждое афферентное волокно устанавливает синаптические контакты со многими мотонейронами. Установлено, что одно волокно, идущее от рецептора растяжения, образует синапсы почти со всеми мотонейронами данной мышцы.
Терминали чувствительных афферентных волокон устанавливают синаптические контакты не только с дендритами и телами иннервируемых ими нейронов, но и сами получают сигнализацию от вставочных нейронов, включенных в цепь пресинаптического торможения, или от аксонов некоторых нисходящих систем.
ПРОВОДЯЩИЕ ПУТИ СПИННОГО МОЗГА
Как уже отмечалось, в спинном мозге имеется целый ряд нейронов, дающих начало длинным восходящим путям к различным структурам головного мозга. В спинной мозг поступает и большое количество нисходящих трактов, образованных аксонами нервных клеток, локализующихся в коре больших полушарий, в среднем и продолговатом мозге. Все эти проекции наряду с путями, связывающими клетки различных спинальных сегментов, образуют систему проводящих путей, сформированных в виде белого вещества, где каждый тракт занимает вполне определенное положение.
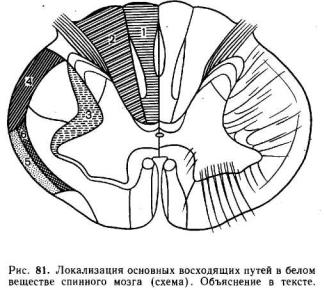
Основные восходящие пути спинного мозга показаны на рис. 81 и в табл. 4. Часть из них представляет собой идущие без перерыва волокна первичных афферентных (чувствительных) нейронов. Эти волокна — тонкий
(пучок Голля) и клиновидный (пучок Бурдаха) пучки идут в составе дорсальных канатиков белого вещества и заканчиваются в продолговатом мозге возле нейронных релейных ядер, называемых ядрами дорсального канатика, или ядрами Голля и Бурдаха. Волокна дорсального канатика являются проводниками кожно-механической чувствительности.

Остальные восходящие пути начинаются от нейронов, расположенных в сером веществе спинного мозга. Поскольку эти нейроны получают синаптические входы от первичных афферентных нейронов, их принято обозначать нейронами второго порядка, или вторичными афферентными нейронами. Основная масса волокон от вторичных афферентных нейронов проходит в составе латерального канатика белого вещества. Здесь расположен спиноталамический путь. Аксоны спиноталамических нейронов совершают перекрест и доходят не прерываясь через продолговатый и средний мозг до таламических ядер, где они образуют синапсы с нейронами таламуса. По спиноталамическим путям поступает импульсация от кожных рецепторов.
В латеральных канатиках проходят волокна спинно-мозжечковых трактов, дорсального и вентрального, проводящие в кору мозжечка импульсацию от кожных и мышечных рецепторов.
В составе латерального канатика идут и волокна спиноцервикального тракта, окончание которых образуют синапсы с релейными нейронами шейного отдела спинного мозга — нейронами
118
цервикального ядра. После переключения в цервикальном ядре этот путь направляется в мозжечок и ядра ствола.
Путь болевой чувствительности локализуется в вентральных столбах белого вещества. Кроме того, в задних, боковых и передних столбах проходят собственные проводящие пути спинного мозга, обеспечивающие интеграцию функций и рефлекторную деятельность его центров.
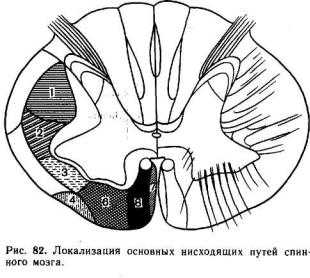
Нисходящие пути спинного мозга также разделяются на несколько самостоятельных трактов, занимающих определенное положение в латеральных и вентральных канатиках белого вещества (рис. 82).
Эволюционно более древние нисходящие пути берут начало от нейронов, ядра которых расположены в пределах продолговатого мозга и моста. Это ретикулоспинальный и вестибулоспинальный тракты. Ретикулоспинальный тракт образован аксонами нейронов ретикулярной формации заднего мозга.
Ретикулоспинальные волокна идут в составе латеральных и вентральных канатиков спинного мозга и заканчиваются на многих нейронах серого вещества, в том числе на а- и у-мотонейронах. Сходную локализацию имеют волокна вестибулоспинального тракта, являющиеся главным образом аксонами нейронов латерального вестибулярного ядра, или ядра Дейтерса. Оба эти тракта не перекрещиваются.
Эволюционно более молодым нисходящим путем является руброспинальный тракт, достигающий наибольшего развития только у млекопитающих. Руброспинальные волокна являются аксонами нейронов красного ядра, расположенного в среднем мозге. Руброспинальный тракт совершает перекрест и идет в составе латеральных канатиков белого вещества.
Окончания руброспинальных волокон занимают в сером веществе спинного мозга более дорсальное положение, чем окончание волокон ретикуло- и вестибулоспинального трактов. Тем не менее часть из этих волокон образует синапсы непосредственно на мотонейронах.
Наиболее важный нисходящий путь — кортико-спинальный, или пирамидный,
тракт, нейроны которого расположены в двигательной зоне больших полушарий. Пирамидный тракт является эволюционно самым молодым. Он появляется только у млекопитающих и наиболее развит у приматов и человека. Волокна пирамидного тракта совершают перекрест и идут в составе дорсолатеральных канатиков над руброспинальным трактом. Окончания кортико-спинальных волокон обнаруживаются главным образом на вставочных нейронах спинного мозга. Пирамидные аксоны, устанавливающие прямые связи с мотонейронами, относятся к миелинизированным волокнам большого диаметра и проводят импульсы с высокой скоростью.
РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ СПИННОГО МОЗГА
В спинном мозге замыкается огромное количество рефлекторных дуг, с помощью которых регулируются как соматические, так и вегетативные функции организма. К числу наиболее простых рефлекторных реакций относятся сухожильные рефлексы
119
и рефлексы растяжения, вызываемые раздражением рецепторов растяжения той же мышцы, которая развивает рефлекторное сокращение. Центральные окончания афферентных волокон от рецепторов растяжения образуют синапсы непосредственно на мотонейронах без дополнительных переключений на вставочных нейронах. Таким образом, дуга этих рефлексов может иметь моносинаптический характер. Указанное обстоятельство, а также высокая скорость проведения по афферентным волокнам, идущим от мышечных рецепторов и по аксонам α-мотонейронов, обеспечивают короткое время рефлекса (что особенно демонстративно в случае сухожильных рефлексов).
Сухожильные рефлексы легко вызываются с помощью короткого удара по сухожилию и имеют важное диагностическое значение в неврологической практике. Рефлекторная реакция проявляется в виде резкого сокращения мышцы. Особенно выражены сухожильные рефлексы в мышцах разгибателей ноги, таких, как четырехглавая мышца бедра (коленный рефлекс) или трехглавая мышца голени (ахиллов рефлекс). Однако сухожильные рефлексы вызываются и в мышцах-сгибателях. На руке они четко проявля-
ются на двуглавой и трехглавой мышцах, на лице — на мышцах нижней челюсти.
Быстрота мышечного сокращения и отсутствие последействия обусловлены способом вызывания сухожильного рефлекса. Адекватным раздражителем для соответствующих рецепторов является
растяжение, мышцы. Постукивание по сухожилию растягивает мышцу только на очень краткий срок.
Зато при этом чувствительные к растяжению рецепторы активируются с высокой степенью синхронности.
Поскольку афферентные волокна, идущие в спинной мозг от рецепторов растяжения, представляют собой довольно гомогенную группу по диаметру и скоростям проведения, афферентные импульсы поступают к мотонейронам в виде синхронной волны. В результате мотонейроны отвечают с незначительной временной дисперсией, посылая в двигательный нерв синхронный разряд, вызывающий короткое мышечное подергивание, сходное с ответом мышцы на одиночное электрическое раздражение двигательного нерва.
Совсем иначе характеризуется рефлекс растяжения, возникающий при адекватном раздражении тех же самых мышечных рецепторов. Естественные растяжения обычно прикладываются к мышцам под действием силы тяжести. Так, при стоянии четырехглавая мышца бедра подвергается растяжению из-за тенденции колена сгибаться под влиянием гравитационных сил. Возникающая в ответ на это растяжение афферентная импульсация характеризуется значительной асинхронностью, так как многочисленные рецепторы растяжения под влиянием постоянной нагрузки генерируют ритмические импульсы, частота которых определяется индивидуальным порогом каждого рецептора. Мотонейроны получают длительные асинхронные импульсы и сами разряжаются асинхронно. В результате этого мышца отвечает плавным длительным сокращением, автоматически противодействующим силе тяжести. Это определяет большое физиологическое значение рефлекса растяжения как механизма поддержания выпрямленной позы или стояния.
Более сложно организованы рефлекторные ответы, выражающиеся в координиро ванном сгибании или разгибании мышц конечности. Сгибателъные рефлексы направлены на избежание различнь\х повреждающих воздействий. Поэтому рецептивное поле сгибательного рефлекса достаточно сложно и включает различные рецепторные образо вания и различные по скорости проведения афферентные пути.
Сгибательный рефлекс возникает при раздражении болевых рецепторов кожи, мышц и внутренних органов. Вовлекаемые при этих раздражениях афферентные волокна имеют широкий спектр скоростей проведения — от миелинизированных волокон группы А До немиелинизированных волокон группы С. Все разнообразные афферентные волокна, импульсация по которым приводит к развитию сгибательного рефлекса, объединяют под названием афферентов сгибательного рефлекса.
Центральное время сгибательного рефлекса довольно продолжительно, что обусловлено его полисинаптическим характером. Сгибательные рефлексы отличаются от собственных рефлексов мышц — миостатических и сухожильных не только большим числом синаптических переключений на пути к мотонейронам, но и вовлечением ряда мышц, координированное сокращение которых обусловливает движение целой конечности. Одновременно с возбуждением мотонейронов, иннервирующих мышцы-сгибатели, происходит реципрокное торможение мотонейронов мышц-разгибателей.
120